
The retinas of the eyes are very complex structures, both across their surface and in depth. Man-made microcircuits are only approaching this level of complexity at the current time. However, they still lack some of the features of the retina , particularly with regard to the fovea, or central region of the retina. The detailed discussion of the features of the retina will be found in Chapters 3 and 4 of PPOCESSES IN BIOLOGICAL VISION. This webpage will address the following topics:
SETTING THE STAGE for the discussion.A particularly unique characteristic of the retina, often overlooked in the literature, is its spherical shape. It is this shape, combined with the variable focal length optical system of the cornea and lens (the lens group) that provide both the extremely wide field of view of the eye combined with its superior acuity along the line of fixation.
The fovea has traditionally been defined using morphological criteria. It looks different from the rest of the retina. There is a more important electrophysiological criteria. The core of the fovea, called the foveola, is connected to the brain by a distinctly different signal path than the rest of the retina. This analytical signal path is associated with the analytical tasks so highly developed in man and only approached by other chordates, even the other primates. This analytical signal path has not been adequately defined and reported in the literature. The remainder of the retina is connected to the brain via the awareness signal path. This is the path normally discussed in the literature.
Man's facility for reading is intimately associated with the analytical signal path. This path leads from the foveola via the pretectum of the midbrain to area 7 of the cortex, also known as the analytical visual cortex. The awareness signal path via the Lateral Geniculate Nuclei to the so-called Primary Visual Cortex plays virtually no role in the higher analytical functions such as reading.
The detailed mmorphological description of the retina is the subject of Chapter 3 of PROCESSES IN BIOLOGICAL VISION. The detailed electrophysiology of the retina is the subject of Chapter 12. The operation of the retina in conjunction with the rest of the visual system is developed in Chapter 11 and 15. The pointing of the retina as part of the optical system of the eye, is described in Chapter 7.3. All of these chapters are available for downloading in beta-release.
PROCESSES IN BIOLOGICAL VISION has defined a large and largely new set of performance descriptors applicable to the visual process in all animals. Some of these descriptors provide a new foundation for many of the previously defined empirical descriptors. It is important to rely upon a consistent set of terms in discussing these descriptors. The following list has been prepared to support the oculomotor generated motions of the eyes. The list is drawn from Section 7.3 of the complete text. It is also presented in support of the web pages related to the reading facility of humans. Further information and references concerning the operation of the oculomotor system of vision can be found in Chapter 7 of the text.
Figure 7.5.3-1 illustrates the size of characters projected onto the foveola and fovea as a function of their size. The black bar represents the diameter of the foveola, a total of 1.2 degrees. The gray bar illustrates the radius of the fovea, a total of 6.2 degrees in diameter. The crosshatched box represents a "square" foveola for purposes of discussion, it is generally circular in form. The letters are drawn at three scales, the size normally associated with good, 20/20 vision, twice that size and four times that size. The spaces between character groups are indicated by the open boxes. Note the number of characters that can be included within the diamerer of the foveola at the various sizes.

The characters are shown in a block font san serif (without any small cusps at the corners). Adding serifs to the characters adds considerably to the legibility of characters because they compensate for the limited resolution capability of the lens group of the eye. However, they complicate standardized testing considerably. It is also important to note that during acuity testing, the letters are usually well spaced. It is much harder to identify individual letters at the limit of our visual performance if they are pushed together as in normal text.
For orientation purposes, the largest characters, labeled 20/80 correspond to 6.28 point type presented at 15 (38 cm.) inches from the eyes, a typical reading distance. This is difficult type for most people to read. They would prefer 9-10 point type. 30 minute high characters corresponds to 20/120 acuity and is represented by 9.4 point type at 15 inches.
At the 20/80 size, it is clear that only about four characters can be imaged on the foveola at one time. Any additional characters spill over into the surrounding fovea, both ahead of and behind the characters currently available within the foveola for perception and interpretation of their meaning. This is important if the signal path of associated with the foveola is distinct from thatn associated with the fovea. This distinction is well documented clinically. Figure 2.6.1-2 of the text illustrates those findings. The reading literature contains a number of discussion concerning how the visual system considers characters visible within the perimeter of the fovea in the reading process. This figure would suggest these peripheral characters play a different and more circumscribed role than found in those discussion. While the visual system is aware of the characters (and the associated spaces)outside of the foveola, and can use these for steering subsequent saccades, these characters cannot be fully perceived and interpreted by the analytical signal channel.
During normal reading, the analytical signal path associated with the nominal foveola can process only three or four characters in a single line of text at one time. This is the reason for the eyes prominent characteristic of moving across a line of text in a series of minisaccades. After each minisaccade, it pauses long enough to perceive and interpret the character group imaged on the foveola. It then proceeds to the next character group.
Initially, these character groups of three or four characters were originally described as words, cat, dog, you, mom, dad, etc. As words became more complex, each group became known as a syllable, cat-a-log, mom-bo, etc. This procedure has continued into the highly complex specialty words of today. tel-e-vis-ion, etc. To speed the process of reading as the words have become more complex, the visual system has evolved a series of optimizations in the reading facility. These include an estimation (guessing) technique where the low level interpretation engines of the brain will make a guess at the the meaning of the next syllable and instruct the saccade generator to skip over it. This proceedure is known as a default saccade. If the assumption is shown to be incorrect by subsequent context conflicts, the saccade generator is instructed to return to a place in the text where the text up to that point does not invovle any context conflicts. The resulting saccade is known as a regression saccade.
At the level of the three or four letter character group, each of the characters frequently impacts the interpreted meaning of the group. A character that impacts the meaning of the group is known as a phoneme. Good examples are cat,mat,rat,hat--car,cat, can, cam, cap--mat, met mit, mut, etc. Each of these changes in phoneme result in a different concept being presented to the analytical engines of the cortex for consideration.
Note that in English, every letter of the alphabet is not used in each character position. From an information theory point of view, this fact lowers the information content of each character group slightly but provides a considerable reduction in the error rate associated with the perception and interpretation process.
FIGURE 7.5.3-3 illustrates the saccades of normal reading using the two sentences;
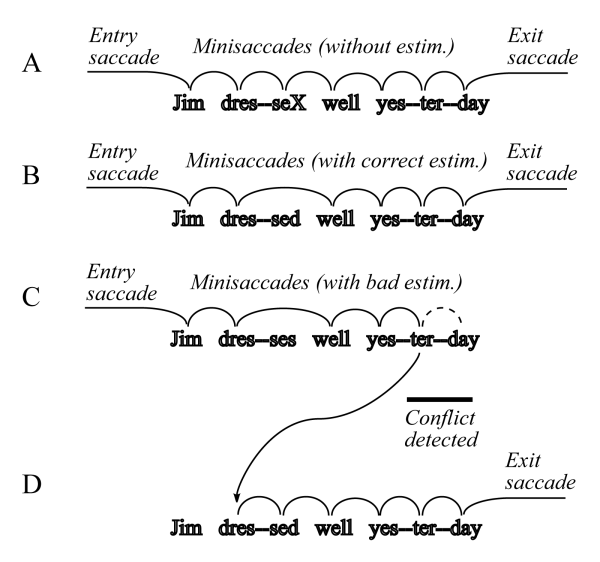
In example A, the system methodically perceived and interpreted each character group in turn. The character labeled X in the character group "sed" would be examined to determine if it was an "s" or a "d." In example B, the system attempted to save time by estimating that the second word was "dressed" based on perceiving only the character group "dre". The total context of the sentence was consistent and the system performed an exit saccade on the way to the next character group. In example C, the system made an estimate that "dre" was part of the word "dresses" and made an immediate default saccade to the first character group of the next word. Perception and interpretation of that character group did not lead to any contextual conflict and the process continued. However, when attempting to interpret the first two or all three character groups of the word yesterday, a significant context conflict arose. This conflict caused the system to abandon the initial saccades sequence that was proceeding and to perform a regression saccade back to the word dressed for reinterpretation.
To understand how the visual system perceives and then interprets the image projected on the retina, it is important to visualize the size of these images versus the size of the individual photoreceptor cells comprising the retina.
Figure 7.5.3-5 illustrates the projection of the word "PRESS" on the human foveola at the scale of 20/20 acuity. Each character is reduced from an angular height of 5 minutes of arc to 3.125 minutes of arc by Snell's Law when projected onto the retina. Under this condition, each character is seen to have a height and breadth equal to approximately 15 photoreceptor cell diameters. (The photoreceptor cells are shown as small circles in a square matrix as opposed to the actual hexagonal packing for simplicity.) The width of each stroke of each character is seen to equal approximately three photoreceptors.
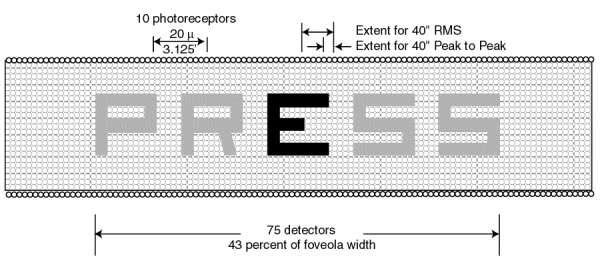
Shown at the upper right is the nominal size of the horizontal microsaccades (tremor) associated with the visual system. The vertical microsaccades are nominally of the same amplitude. Two scales are shown because of the lack of precision in the analyses accompanying the experimental literature. In the worst case, the nominal amplitude of the microsaccades is only two photoreceptor diameters for the 20/20 case. However, even this amount of motion is more than sufficient to scan across the edge of each stroke of each character.
The impact of this relative motion between the projected image of a word or object and the photoreceptors of the foveola and retina is the key mechanism in perception and interpretation of both imagery and text. This mechanism is explored more fully in Section 7.5 of PROCESS IN BIOLOGICAL VISION and in the web page on saccades.