
A major problem in previous discussions of color has been the problem of metamerism. Many sources in object space with different spectral distributions can appear chromatically identical to the human eye. These scenes are called metamers.
Two definitions are important in discussing metamers, that of metamers of course and also of color.
The above can be considered the formal definition of color. It is based on perception. An alternate definition is frequently useful that describes the color of a structure-free field of view in object space that generates the above perception. This definition of color is frequently described as psychophysical color.
Both of the above definitions of color play a role in current colorimetry. However, it will be shown below that it is only the definition based on perception that is precise. Many pairs of psychophysical metamers do not in fact appear to be metamers to the human eye. The differences are frequently significant.
metamers have traditionally been defined in the psychophysical context and is the only context discussed in the colorimetry chapter of Wyszecki & Stiles. However, the fact that two different structure-free fields of view with different tristimulus values frequently appear to be perceptual metamers is troubling. As a result, this work differentiates between the two definitions of Wyszecki & Stiles that they considered equivalent.
Wyszecki & Stiles explored the subject of psychophysical metamers in great detail (38 pages). Whereas the data they summarized is useful, the mathematical analyses are less useful. They attempted to explain the phenomena using the CIE concepts of color space and tristimulus values (based on linearity and additive color). The result is a definition of metameric color stimuli unrelated to biological vision. This definition required that two metamers must exhibit equality in three equations, one related to the tristimulus value r-bar, one related to g-bar and one for b-bar. Thornton has shown that colors defined in this way are not in fact perceptual metamers (Section 17.2.8).
Adopting the actual model of biological color vision, the situation is simpler and more precise. Instead of using the tristimulus values of an imaginary "Standard Observer," the actual absorption characteristic of each chromophore of biological vision is used. Omitting any discussion of the O-channel in human vision, three equations are required to demonstrate a complete metameric match between two color stimuli. However, they are not the three equations found in psychophysical colorimetry. Equation One equates the P-channel values for the two metamers. Equation Two equates the Q-channel value for the two metamers. Equation Three equates the R-channel values for the two metamers. These equations allow for a much larger set of metamers and a much more precise match than does the tristimulus formulation. This range of matches can be subdivided into three distinct classes, the first requiring a precise match in each of the P, Q & R values of the color stimuli, the second requiring a complete match of two ensembles of P, Q & R values and the third requiring a chromatic match of only the individual P and Q values.
While precise metameric matches can be calculated, it is not possible to confirm the uniqueness of such precise matches perceptually at this time. As far as is known, the brain only asserts a complete match based on the somewhat more tolerant ensemble values of P, Q & R. Figure 17.1.2-1 shows the experimental environment associated with chromatic and complete metameric matches. The simpler chromatic match shown in frame (A), typically uses the light reflected by two color samples from a single source of illumination. Because of the interplay of the radiation spectra of the source and the reflectance spectra of the samples, such chromatic matches are a function of the source characteristics. Besides the spectral distribution of the samples in chromatic matches, the match also depends on the average reflectance of the samples used. As a result, the chromatic match equating the P and the Q values may not result in equal R values. Experiments are currently under way to resolve the differences in average reflectance between the currently distributed Munsell Color Atlas and the recently developed comparable Japanese atlas. Frame (B) shows the test configuration for achieving a complete metameric match. By using two separate illumination sources of variable intensity, a match may be obtained that equates the individual P, Q & R values. When obtained, the match is based on the radiant spectral characteristics of the sources and the average reflectance of the samples as well as the reflectance spectra of the samples.
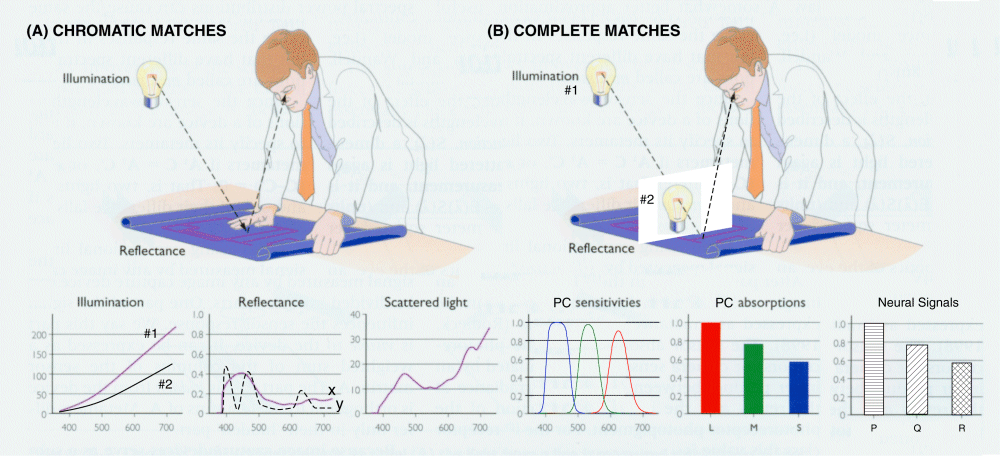
The functions shown in the lower set of frames suggest the parameters that can vary and that must be controlled in these two types of experiments. If two sources are employed, both their intensities and radiant spectra must be controlled or known. The reflectance of the two samples can be significantly different. Scattered light must be minimized for accurate comparisons. The absorption spectra of the actual photoreceptors must be used, and not some arbitrarily transformed set of spectra. While the resulting signal levels at the axons of the spectrally diverse photoreceptors may be of interest, it is the signals resulting from signal processing within the neural section of the retina that are critical to the metameric experiment. It is these signals that are evaluated by the brain in determining a match.
Several second order caveats apply to performing successful metameric matches. Because of the change in the spectral sensitivity of the visual system with intensity of the color stimuli, the experiments should be carried out within the photopic regime, and more precisely the regime of color constancy. To avoid inaccurate results, it is also necessary to carefully define the test protocol used. The most successful tests require a bipartite field with the match determined by concentrating the point of fixation of vision on the midpoint of the bisecting line of the bipartite field. To avoid introducing ambiguities due to Maxwell's spot (Section xxx), it is advisable that the bipartite field have a diameter of less than 1.2 degrees, or much larger than three degrees. Large fields of ten degrees are commonly used. The area surrounding the test samples will affect the state of adaptation, and therefore the color constancy, of the eyes of the evaluator. This area is best made a neutral color not significantly different in illuminance from that of the samples.
This page is in beta release. The author welcomes and will respond to any comments or suggestions left at the comment page. Section numbers of the main manuscript, available on the web, are shown in brackets. The manuscript can provide more detail when desired. The first number shown is the chapter number; it is followed by the section numbers. Download individual chapters.
1Wyszecki, G. & Stiles, W. (1982) Color Science, 2nd Ed. NY: Wiley & Sons
2