
The goal of this PART D is to assemble and interpret the transfer functions of each circuit and stage of the visual process. By concatenating these individual transfer functions up to a given point, a cumulative transfer function can be derived. This cumulative transfer function describes the output signal expected at that point due to any input illumination and describes the input signal conditions applied to any subsequent stage. With the concatenated transfer function available at any point in the system, it is also possible to derive the transfer function of any intermediate section of the visual system. This transfer function can be used to predict the theoretical performance of any experiment that would otherwise depend on a "floating model" for its interpretation.
Finally, with all of the concatenated transfer functions available, it is possible to provide a theoretical foundation for a series of other experiments involving complex signal waveforms. The most obvious of these is the conventional electroretinogram. This work allows the obvious structural differences in electroretinograms to be explained.
With a complete transfer function for the entire visual system available, up to the decoding and presentation of the signals to the brain at the end of stage 3 (see below), it is possible to formulate most of the theoretical characteristics of the visual system and compare them to the observed characteristics determined by psychophysical methods. It is also possible to provide a more structured discussion of systemic failures in the visual system, such as loss in color or spatial perception, based on the complete transfer function of the process. Finally, with the complete transfer function available, it is also possible to explain many unusual visual effects, observed in real life and in the laboratory, in a more structured manner. The definition and illustration of these descriptors of the visual system will be developed in PART E.
To achieve the above level of definition within the circuits, stages and mechanisms of the visual system, it is necessary to employ a highly disciplined approach to modeling of the system. The modeling of man-made signaling systems is a highly developed field. It will be shown that it is now sufficiently developed to be applied to the visual systems of biology as well.
Chapter 11, Introduction to modeling of biological phenomena, presents a review of the modeling procedure adopted from the modeling of man-made systems and shows how they can be applied to the visual system. The chapter opens with an extensive section on terminology and concepts since many of these may not be familiar to the reader. The next major section develops a framework for two aspects of the visual system. The first aspect addresses the correlation of the functional characteristics of the different neuron types with their known morphology (to the extent possible). The second deals with a codification of the various waveforms related to vision that have been previously identified in the laboratory. These frameworks will greatly simplify the discussion to follow. The remainder of the chapter concentrates on evolving two fundamental tools of system modeling. The first is a complete high level block diagram of the visual system that is compatible with all of the known characteristics of that system (particularly with regard to the more extensively understood system of humans). The second is the development of a high level circuit diagram compatible with the above block diagram but introducing more detail relative to the types of circuits involved and the signal path geometries. After this stage, the model and circuit diagram can be further elucidated by outlining the requirements on individual circuits in order to obtain the major overall characteristics of the visual system. This step is divided into three ever more definitive phases. First, the requirements based on the correct handling of large signals are explored. This is followed by the definition of simpler circuit analogs adequate for handling medium and small signals. It is noted that most laboratory experiments involve large signal conditions. The chapter concludes with a short discussion of how the resulting comprehensive models differ from others in the literature.
The above process of documenting the functional details of the visual system is only possible based on the scope of both the preceding analyses in this work and the extensive database in the literature.
As part of this process, a model of the visual system will be proposed that is divided into four major areas labeled stages. Each stage will be analyzed and interpreted from a number of viewpoints frequently of interest to the experimental scientist and the clinician.
Stage 1 encompasses the signal detection process subsequent to the imaging of light onto the retina. This stage will end with the presentation of the signals at the pedicels of the photoreceptor cells.
Stage 2 consists of the signal processing and manipulation carried out between the pedicels and the input terminals of the Activas within the ganglion cells of the retina. Being a functional element in the signal processing stage, the synapses associated with the pedicels will be considered part of stage 2. Similarly, the neurites of the ganglion cells are also key players in the signal processing of stage 2.
Stage 3 encompasses the signal projection circuits of the visual system. The signal projection function is highly developed and easily characterized in Chordata. It is used wherever it is necessary to transmit neural signals over significant distances (typically a millimeter or more). This includes both the efferent and afferent signal paths of the peripheral nervous system (PNS) and the paths within the brain, or central nervous system, (CNS).
Within the visual system, the projection function is used routinely. It is used to transmit signals over the optic nerve to both the LGN and the pretectum. It also used to transmit signals from the LGN to area 17 of the cortex and to transmit signals from the pretectum to area 5 of the cortex. Within the cortex, projection neurons are used to interconnect the different feature extraction engines and logic units of the brain with the initial motor system command generation centers.
In the nominal case, the projection function extends from the input terminal of the Activa within each ganglion cell to the output pedicel of each stellate cell within the brain that receives enervation from a neuron of the optic nerve. Note: the dendritic structure of the ganglion cell is looked upon as part of the signal manipulation function of stage 2, and not as part of the signal projection function of stage 3. Such a division highlights the role of ganglion and stellate cells as transition devices between analog and binary (pulse) signaling.
The further subdivision of stage 3 to accommodate this situation will be addressed in Chapter 14.
Stage 4 consists of all of the higher level signal manipulation and processing that occurs subsequent to the arrival of signals at the brain from stage 3. It may be useful to subdivide this stage to clarify the fundamental difference in the signal manipulation occurring in the Pretectum from that occurring in the LGN and that occurring in the cerebral cortex.
Stage 5 consists of command signal generation within the cortex and controlled by the midbrain acting as gatekeeper. This stage comprises the initial efferent signaling generating engines associated with the signals emanating from the cortex and associated with the visual system (such as the oculomotor system and some aspects of the skeletal motor system). As in stage 4 above, it is difficult to make clear distinctions at this level between the portions of the stage that involve analog signal processing and pulse signal projection within and between areas of the cortex.
Stage 6 consists of the command signal projection paths beyond the cortex.
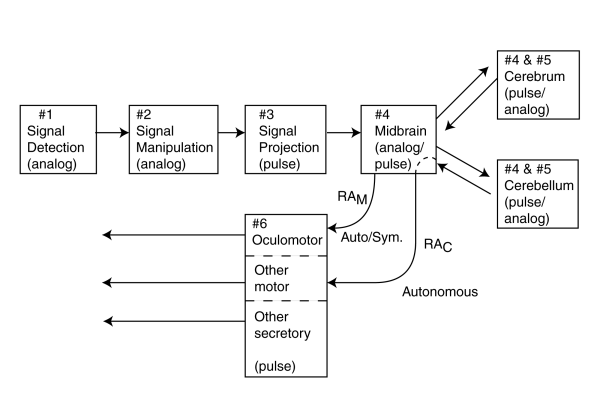
With the identification of the above six stages in a comprehensive model, an additional challenge appears. It is now possible to close some of the loops that include the cortex. It is possible to discuss in detail some of the signals from the above stages as they are turned around, either in the thalamus/mid-brain area or in the cortex, and transformed from afferent signals of the sensory neuron system into efferent signals associated with the motor neuron system. This process can be considered similar to the reflex arcs defined within the peripheral neural system. However, they employ high level signal manipulation and are more similar to sophisticated pulse mode servomechanisms. Extending that notation, the process of turning the loops within the brain will be labeled RAM for those turned at the midbrain and and RAC for those turned in the cortex. The other nomenclature will be discussed in Chapter 11.
Finally, during the development of the model, many details of the operation of the cortex have been illuminated. They have shed new light on the architecture of the cortex and may call for further subdivision of stage 4. This will be discussed more fully in Chapter 15.
It may also be useful to define a subpart of stage 4 to accommodate the discussion of the role of the Superior Colliculus in the operation of the servo system of the visual process. This servo system receives signals from the neurosensory system concerning the line of sight of the eye and delivers signals to the oculomotor system to control this same line of sight.
Chapter 12, Primary Signal Processing--Signal Detection, presents a detailed analysis and description of the photodetection process divided into the role played by the Outer segments of the photoreceptor cells and the role played by the remainder of the cell. The situation is more complex than presented in the current literature and three separate mechanisms are delineated. The first mechanism deals with the photoexcitation/de-excitation of the chromophores of vision, sometimes labeled transduction. This mechanism involves and must be described using quantum mechanical concepts. The second mechanism deals with the translation of the signal into an electrical signal. This mechanism involves quantum mechanical mechanisms, semiconductor physics and conventional electronic circuits with a twist. The output of the first electrical circuit of the photoreceptor cell involves a special avalanche breakdown phenomenon that provides the characteristic of adaptation. This circuit is therefore named the adaptation amplifier circuit. The photoreceptor cell contains a second electronic amplifier circuit. This amplifier acts primarily as an impedance reducing circuit as found in conventional distribution amplifiers expected to support a large number of connections to following circuits. There is a unique feature to this circuit however in that the output circuit load impedance is used as a current to voltage converter. This is a critical function in the overall operation of the visual system. It also introduces a critical difference between the generator waveform measured in the IPM and the generator waveform measured at the pedicel of the photoreceptor cell. The pedicel variant of the signal is a voltage that is the logarithm of the current generating the IPM variant. Several transfer functions for this stage are provided that apply under various excitation conditions.
Chapter 13, Secondary Signal Processing--Signal Manipulation, develops the detailed nature of the various parallel signaling channels found in the visual system.
There are three crucial realizations concerning the initial signal manipulation process. First is that the luminance channel consists of a single channel that involves the logarithmic summation of the current signals from each spectrally distinct photoreceptor channel. Second is that there are three (only two for long wavelength trichromats such as the human) orthogonal chrominance channels in the animal visual system and these involve the logarithmic differencing of pairs of signals from among the spectrally distinct photoreceptor channel. The third is that all of this signal manipulation occurs after the signal level within the perceptual channels of vision have been normalized to an average quiescent level by the adaptation amplifiers. Signals in these channels, although large on average, do not exhibit the large intensity changes found in object (stimulus) space.
The chapter also demonstrates an additional crucial realization. The visual system, and the neural system in general, of animals does not employ negative external feedback. It does employ a more sophisticated form of feedback within a single amplifier circuit. This type of feedback is known as internal feedback. Furthermore, all of the signal processing in stage 1 and stage 2 involves analog signals. In relation to analog signals, the term inhibitory is not well chosen. A signal may be introduced in such a way as to subtract from another signal but the first signal normally cannot inhibit the second signal completely.
This chapter employs certain simplifications in the discussion of the actual signaling paths of vision for purposes of clarity that are explained in the text. Otherwise, a great deal more detail would be required in the discussion. The primary device is the definition of a "straight through" signal path. After this path is defined, a variety of alternate paths are more easily discussed. These paths include divergent, convergent, feedback, phantom (in the telephone context) and encoding paths.
The presence of three distinct levels of baseband matrixing of signals is developed (again telephony terminology). The first is the logarithmic summation channel matrix to obtain the luminance signal. To a large extent, this matrix accepts signals directly from the pedicels of the photoreceptor cells and develops a large number of simple straight through signaling channels. The second level consists of two distinct variants of a lateral matrix. The first variant accepts signals directly from the pedicels of the photoreceptors and generates the difference signals associated with both chrominance and polarization signals (where used) in animals. The second variant of the lateral matrix accepts luminance channel signals from the first matrix and performs an additional level of signal differencing, primarily in the time domain, to produce spatial encoding of the input image to the retina. These signals are primarily related to the spatial appearance of the incident image but involve a degree of spatial encoding expressed in the time domain. Signal dispersal in time plays a much more significant role in vision than recognized in the literature. These signals are extremely difficult to measure using the current techniques of the electro-physiologist.
Following the generation of all of the primary signals to be delivered to the higher processing centers, luminance, chrominance and appearance, the signals are n-ary encoded primarily by the ganglion cells of the retina which constitute the initial elements of stage 3 processing. N-ary encoding is a form of signal dispersal encoding. This encoding greatly reduces the number of individual neural fibers required in the optic nerve. However, the encoding and resulting signals are deterministic and the original signals can be recovered virtually without degradation by the circuits of stage 4.
It is pointed out in the text that there are a small group of photoreceptors in the foveola (within the overall fovea) that are not matrixed as in the above description. These spectrally varied signals are sent directly to the pretectum of the thalamus/mid-brain over paths identical to those used for luminance channels going to the LGN.
Chapter 14, Tertiary signal processingsignal projection. All of the signal processing in stages 1 and 2, and that to be discussed in stage 4, have involved analog or electrotonic signals. It is only in stage 3, involving signal projection over significant distances that the visual system, and for that matter, the peripheral nervous system, employs action potentials. The encoding methods used to insure the highly reliable delivery of the visual signals over the various signaling channels forms the majority of this chapter. Both monophasic and biphasic encoding mechanisms are used. The biphasic encoding is seen to introduce a considerable amount of the signal distortion, asymmetry, and time delay. These effects are causal to many of the phenomena related to flicker. Some of the reasons for the choice of signal encoding are developed, particularly as they relate to the precision optical system (also known as the auxiliary optical system) within the neural systems of the brain. This is the system that controls the generation of optical tremor so vital to the operation of the visual systems of higher chordates, and some higher molluscs.
The detailed signal manipulation function, signal regeneration, performed by the active circuits known as Nodes of Ranvier are shown to be virtually identical to that of the ganglion cells of the retina. The exact circuits used and the precise waveforms generated by both of these circuits are defined. It is also shown that the stellate cells of the LGN and the cortex are the primary decoding circuits of the signal projection stage. They are responsible for recovering the signal information under a variety of specific conditions.
The nature of the signals and signal encoding mechanisms of vision are compared to those of other image recording and reproduction systems.
Chapter15, Higher Level processing--Perception, and signal (vector) interpretation, completes the description of the block and circuit diagrams of the visual system in animals by reviewing the known performance of the brain with respect to vision. The chapter begins with a major section on terminology, required coordinate systems and a variety of concepts applicable to the understanding of the visual system at the highest level of signal sophistication. Following a discussion of the differences in the higher order elements of the visual systems between phyla, orders and in some cases families, detailed block diagrams of the initial elements of the cortex involved in vision are presented. These stress the architecture of the cortical part of the visual system. This architecture is shown to consist of a large group of "feature extraction engines" operating in a star network configuration. The visual signals are introduced into this architecture at two distinct locations. An argument is made that the so-called primary visual cortex associated with area 17 of the brain is actually the secondary entry point in the higher chordate, and particularly the higher primates. In these animals, it is likely the entry point near areas 5 and/or 7, which receives the higher precision data from the foveola of the eye, is more appropriately labeled the primary visual cortex.
By examining the nature of the signals recorded at different location in the animal brain, of higher primates particularly, the detailed signal topology from the retina to the cortex, at least up through area 7a, can be deciphered. This signal topology embraces a large amount of time dispersal encoding that is not elucidated in the current literature. This time dispersal is of particular importance in the operation of the LGN. Within the cortex, it is shown that the various feature extraction engines of vision operate essentially asynchronously with respect to other engines except on the most gross basis. A feature of the analog signaling environment within an engine is its ability to operate very fast over very short distances. There is no limit such as that associated with the action potentials of the projection system. The outputs of individual engines are required within a matter of milliseconds but their internal "clock speed" may be on the order of microseconds.
This chapter also explores the two major servo loops of vision. The main loop can be described as a sampled data servo system with an analog general purpose central processing unit (CPU) not unlike many man-made servo systems prior to the change over to digital CPUs in the 1970's. This loop involves the cortex. The precision servo loop (incorporating the auxiliary optical system of the neural system) controls the fine motions of the ocular globes and does not involve the cortex. It is also a sampled data type servo loop with an analog special purpose central processing unit. This special purpose based computing system operates in a variety of modes. One of the most important is to accept signals from the vestibular system in order to maintain the desired visual line of fixation in the presence of unrelated movement by the head and skeletal systems.
The subject of the perception of spatial relationships by the feature extraction engines (and the related perception of motion) is not addressed in this chapter. However, the task of feature extraction with regard to luminance and chrominance information is addressed. It is shown that these processes are accomplished in a linear manner. This fact allows the signals recovered by the signal recovery circuits (stellate cells) of the signal projection system, stage 3, to be considered as characterizing the perceptual performance of the visual system. By proper projection back to the retina, it is also possible to consider the signals recorded in the latter circuits of stage 2 to also be considered indicative of the perceptual performance of the visual system.
A short catchall section concludes this chapter.
Return to the website home page