
COLOR CONSTANCY
A Special Topic in
PROCESSES IN BIOLOGICAL VISION
Last Update 02 July 03
Rhodonine™ and Activa™: See Citation Page
INTRODUCTION
The subject of color constancy has become more important recently due to the
interests in robotic vision and electronic cameras. The need has been to
understand color constancy in human vision so similar techniques can be introduced
into these devices.
A variety of discussions of this subject have appeared on the WEB in recent
times. Most of these have lacked any theoretical foundation related to the actual
mechanisms of human vision resulting in color constancy. Most of these have also
failed to define the illumination range over which the proposed mechanism applies.
Based on the recently completed book, PROCESSES IN BIOLOGICAL VISION, the
phenomena of color constancy and the underlying mechanisms causing it can be
defined in considerable detail. The overall phenomena is discussed in
Section 17.4.5 of that work. The underlying mechanisms are discussed in
Chapter 12 and Section 17.1.1.
Contrary to the intimation or claim in some of the literature and WEB papers,
color constancy does not involve cognitive processing or any de-convolution in
the spatial or spectral domains to determine the reflectance coefficient of a surface.
It does involve temporal frequency filtering within the photoreceptor cells of the
eye.
The same techniques used in the visual system to achieve color constancy have
been in use in industrial and military television equipment since the 1950's.
BACKGROUND
Color constancy is closely related to the phenomena known as reciprocity
failure in photographic film technology. In a sense, it is the complement to
reciprocity. In human vision color constancy is directly related to the ability
of the system to work over an extended radiant intensity range. Both phenomena
and the transient known as dark adaptation are all controlled by the adaptation
amplifiers found within each photoreceptor cell of the retina. This amplifier
has been described in detail in the above text.
Some of the terminology used in the following paragraphs may be unfamiliar
to some readers. A short glossary is provided at the end of this paper.
The Individual Adaptation Amplifier
The morphologically defined photoreceptor cell contains three major components;
the Outer Segment, the Inner Segment, and the Neural Segment. Each segment
performs a separate and distinct role in vision. The Outer Segment is not an
internal part of the cell. It is an external structure associated with the
cell and formed by the Inner Segment of the cell by secretion and extrusion.
When coated with the appropriate chromophore of vision, the Outer Segment
becomes a passive transducer of radiant energy into energy of excitation within
the structure. This transduction process is inherently linear.
The energy of excitation is transferred to the neural portion of the cell by
a mechanism very similar to that found in a photo-transistor. This process will
be called translation to differentiate it from the above transduction process.
The translation process is linear in the first order but does involve some
second order non-linearities that do not impact this discussion. The dendritic
structure associated with the Neural segment of each photoreceptor cell is a
distributed, open base, electrolytic semiconductor device known as an Activa.
The Activa is a biological transistor. This device is arranged in a differential
pair type of circuit with a second Activa that acts as a distribution amplifier
and a current to voltage convertor. The open base input Activa
is known as the adaptation amplifier because of its unique performance capabilities.
The adaptation circuit [Sections 8.6, 12.5 & 17.1.1] is unique in three respects:
- Adaptation is controlled by the percentage of the chromophore material associated with all of the disks of a specific photoreceptor that is electronically excited at a given time. This excitation is observable by measuring the degree of bleaching of the retina from its fully dark adapted condition. The excitation level is reduced by the average current flow from the base of the adaptation amplifier into the liquid crystalline chromophore surfaces. The average current flow from the base is a precise fraction of the current through the collector of the adatation amplifier.
- The load impedance in the collector lead has a time constant that is finite,
between 0.6 and 3.0 seconds depending on the species. The power supply supporting that load impedance is
very poorly regulated. It exhibits time constants of 2 minutes and 10 minutes.
- The output signal from the adaptation amplifier is taken from the emitter
terminal.
Because of characteristics 1 and 2, the chromophoric mass associated with the input of the adaptation amplifier exhibits a very large variation in absorption cross-section. This variation is the primary cause of the loss in sensitivity at high stimulus levels.
Because of characteristic 3, the overall circuit exhibits a large amount of negative
internal feedback.
The Individual Distribution Amplifier
The distribution amplifier, also located within the Neural Segment of the
photoreceptor cell, is configured as a "grounded base" amplifier. In this
configuration, it offers a unity current gain between the current at its emitter
and the current at its collector.
The combined amplifiers of the Photoreceptor Cell
The complete circuit diagram of the photoreceptor cell is shown below.
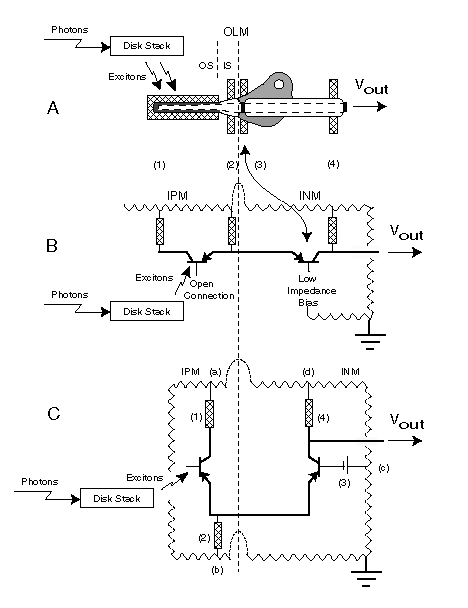
The detailed discussion accompanying this figure is available in
Section 10.10.7. By placing the adaptation
amplifier in a differential pair configuration with the distribution amplifier,
an additional feature is introduced. This configuration is designed to maintain
a constant current through the common emitter(s) to ground load (2). As a result of
this feature and the time constants discussed earlier, a very high degree of negative
internal feedback is introduced into the overall photoreceptor cell circuit at
low frequencies. The average current in the load attached to the distribution
amplifier collector (4) remains essentially constant. At higher frequencies, the
circuit exhibits considerable gain. The effect is to normalize the output
signal DC gain while providing a variable AC gain that can be very large at
low input signal levels. The variation in AC gain is from 1:1 to about 3,500:1
in a typical photoreceptor.
The above variation in gain as a function of input illumination forms the
basic mechanism of both adaptation to changes in input intensity level and to
the phenomenon of color constancy.
The spatial and spectral configuration of the visual system
The human retina consists of a mosaic of three interdigitated arrays of
spectraly selective photoreceptors. The detailed parameters of each of these
arrays are not well documented. However, the effect is to provide a mosaic that
samples each of the the important features of the scene in each of three
separate spectral regions. Each of these samples is converted into an electrical
signal by a separate set of spectrally selective photoreceptor cells.
All spatial signals are converted to temporal signals for transmission from
the retina to the brain. In this process, all fine image detail is converted
to a temporal frequency range that insures that the resulting signal will pass
through the adaptation process successfully without degradation.
The Dynamics of the visual system
To understand the dynamics of the visual system, it is necessary to review
the way in which the photoreceptors in a given location of the retina are
interconnected. A variety of simple symbolic representations of each adaptation
amplifier are possible. The symbolic representation shown for the three adaptation
amplifiers is for illustrative purposes only.
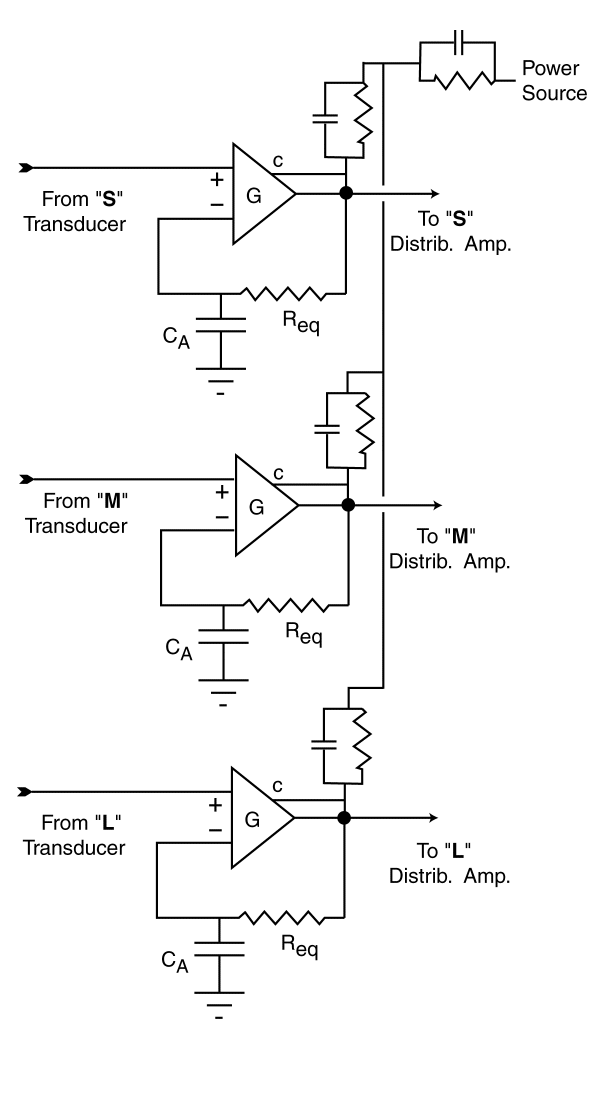 For purposes of this discussion,
the signal from the transducers will be considered a voltage instead of a
current because most people find it easier to think in terms of voltages.
It shows an external feedback loop that does not exist in the actual amplifier.
In this figure, signals are acquired from the same region of the scene imaged
on the retina by the three transducers of the Outer Segments. The signals from
the individual transducers are applied to the individual adaptation amplifiers
as shown.
For purposes of this discussion,
the signal from the transducers will be considered a voltage instead of a
current because most people find it easier to think in terms of voltages.
It shows an external feedback loop that does not exist in the actual amplifier.
In this figure, signals are acquired from the same region of the scene imaged
on the retina by the three transducers of the Outer Segments. The signals from
the individual transducers are applied to the individual adaptation amplifiers
as shown.
The capacitor CA and the impedance Req form a low pass
filter in the feedback path from the output of each amplifier to the negative,
or inverting input on the left of each amplifier. This circuit has the effect
of holding the DC voltage at the output of each amplifier at a constant voltage
regardless of the voltage applied to the input from the transducer. However,
the AC gain between the transducer signal and the signal going to the distribution
amplifier may be quite high under this condition. In the case of vision, the
instantaneous gain of the amplifier is a function of the voltage applied to
the collector (c) of the amplifier. For large increases in the input signal,
the gain of the circuit will decrease. Because of these two distinct processes,
the output signal to the distribution amplifier has a constant average value
and a AC signal that deviates from the average by a constant amount for a given
input contrast. Each spectral channel of the visual system, and all subsequent
perceptual channels operate in this fixed amplitude condition as long as each of the
adaptation amplifiers is within its operating range. This range extends from
a nominal minimum gain of 1:1 up to a maximum of 3500:1 and defines the
photopic region of vision.
Figure 17.1.1-2 of the text is shown here. It discusses the apparent brightness
as a function of illumination level in human vision. The dashed line indicates the
average brightness perceived by the individual from complete darkness to the point
of pain. The remarkably wide flat expanse of this figure is primarily the result
of adaptation within the photoreceptor cells.
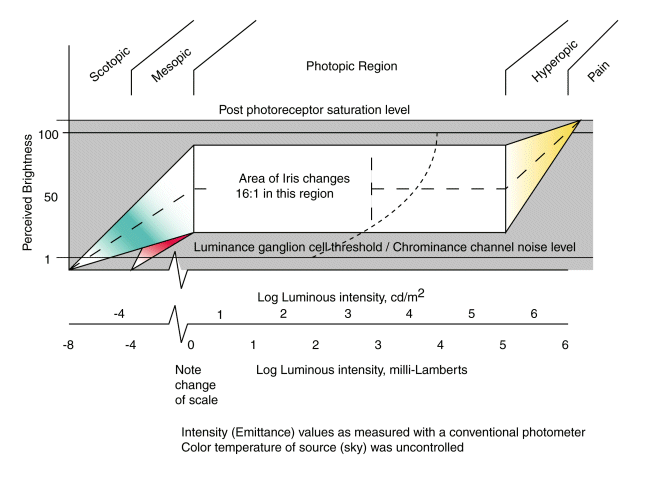
This figure is representative of the situation in the absence of color
changes in the illumination. The color changes shown in the figure are due to
secondary processes within the system. As discussed below in detail, if the color
temperature of the illumination changes, or the reflectance of the scene shows a
predominance in one region of spectral absorption by the photoreceptors, the situation
changes considerably to maintain "Color Constancy."
Initial condition of the adaptation amplifiers
This paragraph is only conceptual. There is not enough data at the current
time to confirm it in detail. However, it is approximately correct. The initial
operating gain of each adaptation amplifier appears to be set as if it expects
the input illumination to have an equal flux per unit wavelength distribution
across the visible spectrum. This condition is produced by a nominal blue sky
absent direct light from the sun, i. e, highly scattered sunlight. This
illumination can be represented by a color temperature of 7053 Kelvin although
it has frequently been described as equal to a color temperature of 6500 Kelvin.
These values are not significantly different absent a specific criteria. It
appears that when the eye is completely dark adapted, it is optimized for the
7053 Kelvin scene spectrum.
Operation of the adaptation amplifiers in unison
When the eye is exposed to a 7053 Kelvin color temperature scene and the
intensity of the scene is varied, the [luminous intensity function] of the eye
is obtained. This function exhibits a broad area of constant perceived brightness
for a variation in input intensity of about five orders of magnitude. This wide
range is accounted for through the operation of the iris and the adaptation
amplifiers. The Iris provides a factor of 16:1 out of this total range and
the adaptation amplifiers operating in unison account for the other factor of 3500:1.
The hyperopic region is defined as that region of illumination higher than these
two mechanisms can accommodate. Two regions are defined below the photopic
region. The mesopic region is an area where all of the adaptation amplifiers
are operating at maximum gain and the iris is fully open. This is an area of
reciprocity failure due to secondary processes in the L-channel, and a
decreasing ability to perceive both color saturation and hue due to signal to
threshold level considerations. It is also an area where the conventional
Principle of Univariance fails in the L-channel. Below this level is the
scotopic level. It is defined by the complete absence of visual perception in
the L-channel of vision relative to the S- and M-channels.
Differential operation of the adaptation amplifiers
Color constancy is present throughout the photopic region but it generally
goes unnoticed until the average luminosity of the scene, over a specific spatial
angle, can no longer be represented by a color temperature of 7053 Kelvin.
When this occurs, each adaptation amplifier attempts to automatically compensate
for the difference in average luminosity level sensed by its transducer. The
result is different gains in the amplifiers of each spectral region of vision.
Under these conditions, the perceived signal levels within the visual system
from the pedicels of the photoreceptors to the higher centers of the cortex
remain constant and the chrominance channels of the visual system continue to
report the same colors relative to the fine detail in the scene (with minor
changes to be discussed below). This effect can be visualized if the conditions
in the scene, the gain parameters of the adaptation amplifier and the perceived
signal levels in the visual system are examined. The following figure illustrates
the perceived variance in brightness versus color temperature.
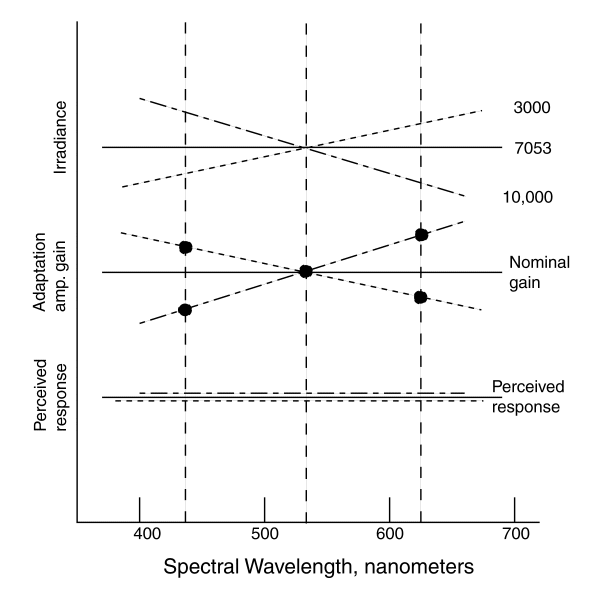
If the average small area radiance of the scene is represented by
a 7053 Kelvin source, the nominal gain of each of the adaptation amplifiers is
the same as indicated by the solid lines in the upper two frames. Under this
condition, the perceived response as a function of wavelength remains a relative
constant as shown in the lower frame. If the color temperature of the radiance
is lowered as suggested by the doted line, the level of irradiance detected by
each transducer is the integral of the absorbed flux. The value of this flux
causes each adaptation amplifier to change its gain to compensate for this change
in irradiance as shown in the middle frame by the dots at the centroidal wavelength
of the chromophores. The dotted line is drawn through these three values merely
for illustration. Following this compensation, the relative perceived signal
levels in the visual system remains at a constant level at the centroidal wavelengths
of the chromophores as shown in the lower frame. The dotted line is only
conceptual, the absolute perceptual level as a function of wavelength is
actually given by the luminous efficiency function. The dash-dot lines of
the three frames show the example of a higher color temperature irradiance.
Combined group and differential adaptation
Group adaptation (related to a change in overall intensity) and differential
adaptation (related to color temperature of the irradiation) can occur
simultaneously as long as all adaptation amplifiers remain within their
individual operating dynamic ranges. The impact of an adaptation amplifier
reaching its maximum gain, and entering its scotopic region, is easily seen
with respect to the S-channel. As the color temperature of the light source
is reduced and the intensity level is also reduced, the S-channel adaptation
amplifier reaches maximum gain first. This condition, typified by artificial
incandescent illumination compared to mid day solar illumination, results in
a lack of perception of blues. (Always illuminate your paintings brightly with
a high color temperature source.)
Machine Vision and Color Constancy
The methods of adaptation used in vision are available for use in robotic and
other man-made imaging systems. In fact, they have been widely used in such
systems for many years. The author implemented such designs during the 1950's
in vidicon based television equipment for the military and industry. He was
awarded a patent on a charge coupled device (CCD) based imaging system in the
1970's that provided automatic exposure control over a wide range. When used
in a multi-CCD array for color imaging, it was quite capable of providing color
constancy over a range of at least 1000:1 without support from an iris. In the
case of the vidicon cameras, both simple resistors and active impedances were
used to achieve the same type of poor or spongy power supplies as used in the biological
systems. In the CCD's, varying the integrated signal in devices containing a gate
designed for exposure control provides the same capability at low cost.
Glossary
AC--A term derived from alternating current and used to describe the portion of an
electrical waveform that changes rapidly with time.
AC coupled--Two circuits that are connected so they only pass AC signals.
AC gain--The gain of an amplifier with respect to rapidly changing signals.
DC--A term derived from direct current and used to describe the portion of an electrical
waveform that does not change rapidly with time.
DC coupled--Two circuits that are connected so they pass both AC and DC signals.
DC gain--The gain of a circuit with respect to slowly changing signals.
Feedback--Refers to a process where at least a portion of a signal is returned
from the output to the input of a circuit.
- External feedback--A process where an external physical signal path extends
from the output to the input of an amplifier.
- Internal feedback--A less obvious form of feedback, typified by a common
impedance element appearing in both the input and output circuits of an
amplifier. There is no explicit feedback path around the amplifier.
Gain--The amplification factor, or ratio of the output signal amplitude to
the input signal amplitude of an amplifier.