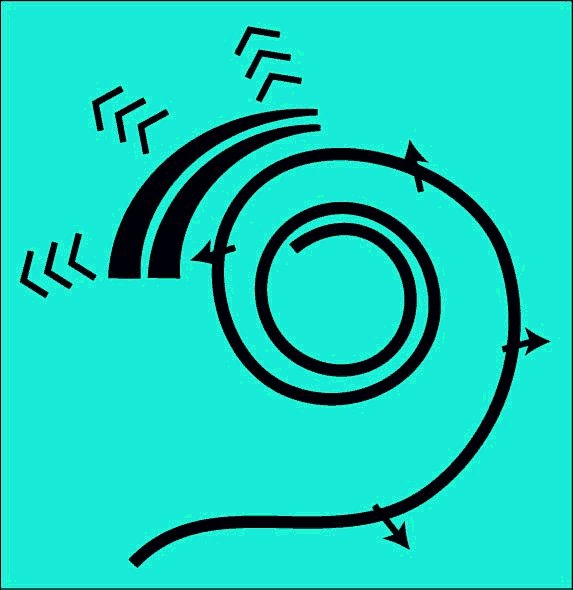
Two distinct dynamic models are required to define the operation of the cochlea. The first describes the motions along the longitudinal length of the cochlea (shown here uncoiled). The energy propagating longitudinally along the cochlea is confined to Hensen's stripe of the cochlear partition. Hensen's stripe is a feature of the gel surface of the tectorial membrane.
Acoustic energy travels along Hensen's stripe as a surface acoustic wave with its amplitude transverse to the surface of the gel. The energy travels at a fixed velocity regardless of frequency. As the energy travels along Hensen's stripe, the energy is separated from Hensen's stripe and directed toward the Outer Hair Cells. The highest frequencies are separated first. As the energy is separated, the envelope of the total energy is degraded until only a sine wave component remains. This component is at the lowest frequency of the applied acoustic energy. This frequency is generally but not necessarily the fundamental frequency of the Fourier series describing the envelope of the applied excitation.
The total acoustic energy traveling along Hensen's stripe at a given point is sensed by the rigid cilia of the IHC and converted into an electrical signal. The conversion efficiency is quite high. However, a small amount of energy is not converted and is seen to cause motion of both the tectorial membrane and the basilar membrane acting as inertial masses.
The model described further below is used in all mammalian hearing. The model is also applicable to the monotremes, the egg-laying but nursing animals such as the platypus and the echidna of australia. A similar mechanism may or may not be used in amphibians, reptiles and birds.
For the scope and top-level description of the SAW-based Electrolytic Theory of Hearing, see page 18 of Chapter 1 of the e-book "Processes of Biological Hearing"
The following material related to the longitudinal structural model of the cochlea is drawn from Section 4.3 of Chapter 4 of "Processes in Biological Hearing." A draft of this material can be accessed from the Home Page of this website.
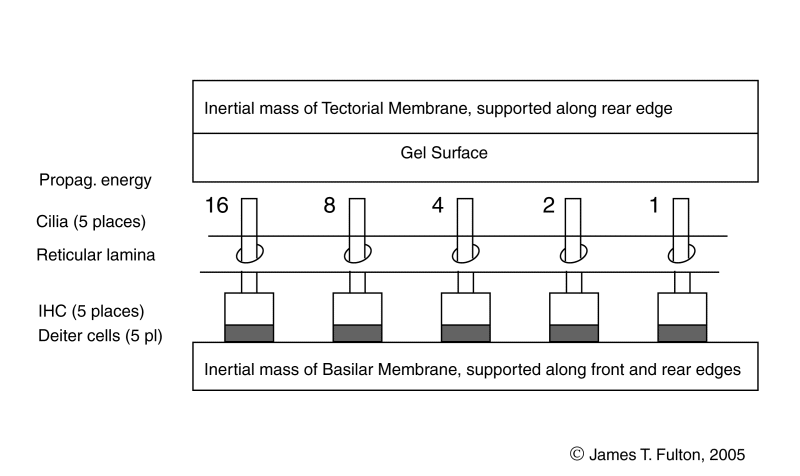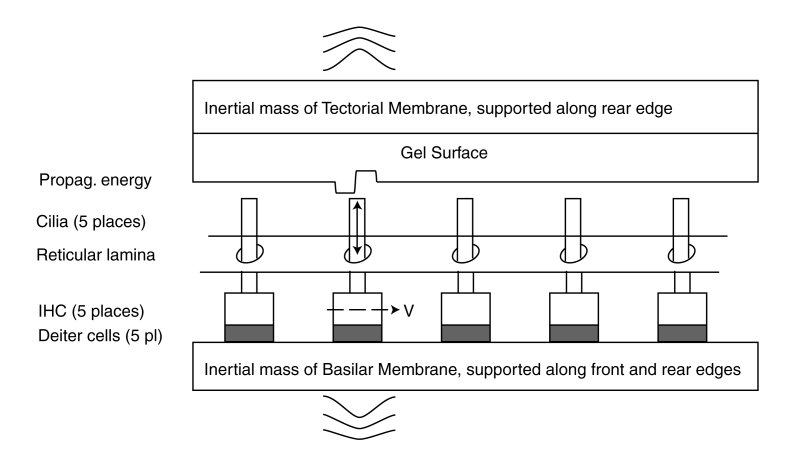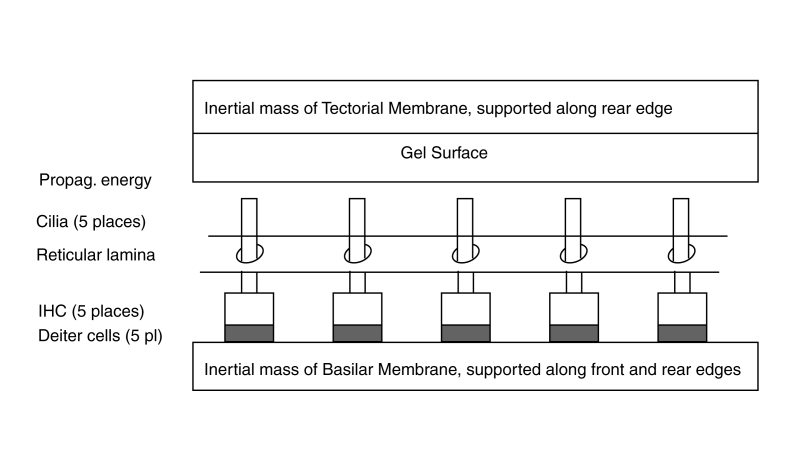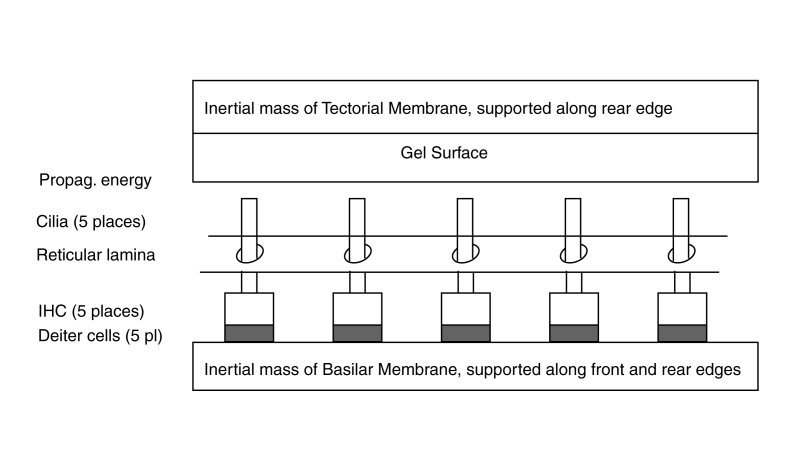
The active portion of the cochlea consists of the elements within the cochlear partition and associated with the Organ of Corti. The dynamic model of the longitudinal cochlear partition consists of many elements. Of particular importance are the elements of the tectorial membrane and the elements mounted on the basilar membrane. Crucially important are the inner gel surface of the tectorial membrane and the Inner Hair Cells of the basilar membrane. These elements are arranged between the bulk of the tectorial membrane and the bulk of the basilar membrane acting as inertial masses. The energy delivered to the Organ of Corti arrives via the Hensen's stripe portion of the gel surface of the tectorial membrane.
Acoustic energy travels along Hensen's stripe as a surface acoustic wave with its amplitude transverse to the surface of the gel. The energy travels at a fixed velocity regardless of frequency. As the energy travels along Hensen's stripe, the energy is separated from Hensen's stripe and directed toward the Outer Hair Cells. The highest frequencies are separated first. As the energy is separated, the envelope of the total energy is degraded until only a sine wave component remains. This component is at the lowest frequency of the applied acoustic energy. This frequency is generally but not necessarily the fundamental frequency of the Fourier series describing the envelope of the applied excitation.
Since the distance between the tectorial membrane and the basilar membrane is essentially fixed, the energy of the surface acoustic wave must be absorbed by the IHC. Becauseof the ratio of the piezoelectric compliance of the IHC and the masses of the tectorial and basilar membranes, a majority of the energy is absorbed by the IHC. Only a residual amount is converted into motion of the two membranes.
The energy absorbed by the piezoelectric portion of the IHC is converted into an electrical voltage that is delivered to the neural system via the synapses of the IHC, a sensory neuron.
The following animation is meant to describe how a square pulse of energy applied to the oval window of the cochlea is processed within the cochlear partition. It is important to note that the cochlea has been uncoiled for illustrative purposes only. The process of extracting individual frequency components from the cochlea requires it be curved.
The first frame of the animation is provided for orientation purposes. Callouts are provided to all of the relavent portions of the cochlear partition. Following this frame, a series of frames are shown describing the movement of the applied pulse down the gel surface of the tectorial membrane and its conversion into an electrical signal.
Five Inner Hair Cells are shown spaced equally along the length of the basilar membrane. Because of the frequency extraction mechanism, this equal spacing describes the location of harmonically related IHC (and the location of the OHC discussed in the lateral dynamic model animation).
If an applied pulse of 1000 Hertz with a duration of one millisecond and a longer repeat interval,the resulting signal will exhibit frequency components spaced at multiples of 1000 Hertz. Under this condition, the IHC can be described at locations along the gel surface corresponding to the 16th, 8th, 4th, 2nd and fundamental frequency of the applied pulse. The IHC are so labeled in the animation. A great many additional IHC are found between each of the IHC shown.
The envelope of the applied energy begins as a high quality square wave. As it progresses, the higher frequency components are removed and the pulse becomes more rounded, ending as a sine wave at the last relavent IHC.
At each IHC, the amplitude of the SAW wave is impressed upon the cilia and transferred to the piezoelectric portion of the individual IHC. A small amount of energy is not absorbed but is transferred to the tectorial and basilar membranes. This residual energy causes motions of these membranes that can be measured. The amplitude of these motions decrease with distance along the Organ of Corti.
The piezoelectric action of each IHC generates a voltage proportional to the envelope of the applied energy. Each signal is delayed by the travel time required by the signal to travel between pairs of IHC. For the propagation velocity of the energy in humans, the output pulses occur at 0.5 millisecond intervals.
Notice the pressure waves generated within the fluids surrounding the tectorial and basilar membranes. The displacement of the two membranes extend to considerable distances along the Organ of Corti to each side of the IHC because of the finite stiffness of these membranes. The resulting pressure waves exhibit similar width.
The amplitude of the tectorial and basilar membrane motions are practically negligible, typically much less than one-thirtieth of the amplitude of the cilia motion. These residual motions are smaller than the thermal motion of individual molecules within these membranes.
Go to the next animation in the dynamic model chain.
Go to the roadmap of available animations of hearing.
Return to the website home page.