
As indicated by the title, this work is intended to describe the vision process in all animals. The intent is to do this at the detailed and mathematically defendable level of the individual mechanism. With this accomplished, a comprehensive discussion can be presented at an equally defendable level of the operation of the overall visual system. This discussion will culminate in the presentation of a new set of performance descriptors defining the visual process at a level of detail never presented before. An approach to this subject based on taxonomy will be limited to a small portion of Chapter 1 where it is appropriate.
The subject matter is broad and the animal kingdom has evolved an unbelievable variety of variations on an underlying theme. Practical compromises must be made. Therefore, in discussing the fundamentals of vision, the characteristics of a variety of animals will be discussed to show how the vision of all animals can be explained within the context of this work. Beyond the fundamentals, the vision of man is both more widely explored and important to the community. Unfortunately, much of the data in the literature has been interpreted based on inadequate, and frequently partial (floating) models. Furthermore, a lack of adequate experimental control has frequently resulted from the lack of a comprehensive model. This work will attempt to provide a framework for an even more thorough understanding of the human visual system and the vast array of psychophysical data available.
The inconsistent use of terminology is a major problem in the literature of the visual sciences. In this work, it has been necessary to define many terms more precisely than in the prior literature in order to provide precise understanding of many concepts. A Glossary is provided at the end of the work to aid in interpreting the precise terminology used.
In this Preface, only a few references have been provided; primarily to support matters that are not discussed further in the main work. It has been necessary to present several ideas in the Preface that are contrary to the general thinking of the visual science community in order to help the reader follow the development of this work. Every attempt has been made to expand on the comments made in this Preface in the main work. That is where detailed references will be found.
When Newton presented his work on Celestial Mechanics, he stopped short of a complete work--treating the planets and the Sun as a series of individual two-body problems. At the time, he is said to have remarked concerning the remaining and unaccounted for perturbations in the orbits of the planets: "The rest is in the hands of the Creator."
Laplace, the great mathematician, set himself the ambitious task of refining and perfecting Newton's calculations involving mechanics in a book that should offer a complete solution of the great mechanical problem presented by the solar system, and bring theory to coincide so closely with observation that empirical equations would no longer find a place in astronomical tables. The result was the highly acclaimed five volume set, Mecanique Celeste.
When Laplace presented the first edition of his work to Napoleon--so the story goes--Napoleon alluded to Newton and remarked, "Monsieur Laplace, they tell me you have written this large book on the system of the universe, and have never even mentioned its Creator." To this Laplace replied bluntly, "I had no need for that hypothesis."(1)
The author, although less esteemed, finds himself in an analogous situation concerning some of the hypotheses that have matured in the field of vision. When asked about why these hypotheses do not appear in this work, the fundamental reply of the author is that: "There was no need for that hypothesis." This statement applies to the following hypotheses:
+ Functionally different photoreceptors, rods and cones, based on morphological differences.
Using only floating models, based primarily on the work of early anatomists. Past investigators have continually attempted to assign a chromatic function to one morphological structure identified in the retina and an achromatic function to a second morphological structure. This has caused untold confusion since 1) the variation in photoreceptor cell morphology is not truly represented by a dichotomy, even of sub-parts of the photoreceptor cells; 2) the proposed achromatic class, the rods, has been shown to exhibit various narrowband chromatic spectra; and 3) no broadband spectrum, required by an achromatic rod, has ever been recorded from a single photoreceptor. The difficulty has been compounded by the failure of the community to extract successfully a single chromophore from a cone that exhibits the correct narrowband chromatic spectra. Merging information from the disciplines of bio-chemistry, dye-chemistry, and signal processing allows the definition of a comprehensive model that explains how the deuterostomic retina operates using only three types of photoreceptors to compute the exact overall achromatic spectrum and the difference signals used to provide the sensation of chromatism.
+ An external cell membrane of the photoreceptor cell that encloses the disk stack known as the Outer Segment.
In the first half of the 20th century, the outer segment of the photoreceptor cell, containing the disk stack, was assumed to be an integral part of the cell and enclosed by the outer cell membrane. In the last half of the century, electron microscope images have failed to show this to be the case; both for single cell images and for images of a detached retina where many outer segments are shown sheared in half, no sign of any debris is seen that could be associated with the tearing asunder of all of the putative external cell membranes. Further more, recent tests show that the photoreceptor cell is a neuro-secretory cell--excreting a protein substance, known colloquially as opsin. This material is coated, after secretion, by the chromophoric material transported to the site from the retinal pigment epithelium (RPE). The chromophoric material is designated here as rhodonine. These observations are all compatible with the comprehensive model presented here.
+ Rhodopsin in two different contexts; where the protein portion of the molecule plays the role of a chromophore, and where the ligand is a non-resonant derivative of Vitamin A.
While rhodopsin has been proposed as the photo receptive material of the retina for a very long time, long before the suffix -in was adopted as a terminus for the name of a protein, the exact nature of this material has never been specified. Limited to the biochemical tools available in the 1930's through 1950's and lacking a comprehensive model of the photodetection process, Wald arbitrarily defined (ordained) the composition of rhodopsin as a combination of a derivative of Vitamin A and a protein, opsin, which provided the chromophoric absorption site. How these elements were combined was open to further investigation and subsequent analysis, primarily intuitive, focused on the use of a Schiff base structure. This overall structure did not exhibit the required spectral response; therefore additional intellectual and mathematical analysis was performed focusing on possible protonation of the Schiff base--without obvious success. As with rods and cones, above, merging information from the disciplines of bio-chemistry and dye-chemistry has produced a more comprehensive model of the photochemistry of vision in all animals; this model has redefined rhodopsin as a more loosely linked combination involving the hydrogen bonding of a liquid crystalline chromatic material to a benign substrate made up of protein. The liquid crystalline material is one of four homologs of a resonant retinoid, named rhodonine, derived from Vitamin A. This homologous series does exhibit the exact spectral absorption properties associated with animal vision, including the ultraviolet spectrum found in insects and other selected animals.
+ Glutamate cascades to account for the amplification of the initial photon generated current.
The glutamate cascade is a recent hypothesis in the signal theory of vision; it came about when the community failed to develop a comprehensive model related to the Photoexcitation/De-excitation (P/D) problem of vision and because of this failed to solve the detailed differential equation applying to the problem. A hypothesis was then proposed, by the same group of investigators who failed to solve the equation, based on a material found to occur (conveniently) near the site of the P/D process. Other investigators have determined that glutamate does not have a role in transduction or neuro-transmission. With an adequate comprehensive model, the solution of the P/D equation, and the definition of the first activa (amplifier) in the signal chain, there is no need for a process such as a glutamate cascade. The cascade was originally defined based on a floating model. An alternate role for glutamate is provided that appears compelling.
+ Cascaded discrete electrical filter stages within the retina to account for the temporal frequency response of electrophysiologically measured waveforms.
Investigators have made electrophysiological measurements for many years suggesting the product of the input stimulus and the transmission path of animal vision resulted in a signal with a variable width temporal frequency spectrum. Without a comprehensive model of the process they were interpreting, they have used a floating model and have attempted to account for this variability by introducing a multiple stage, generally lumped constant, electrical filter into the transmission path. Using a comprehensive model, the solution of the complete Photoexcitation/De-excitation differential equation clearly shows that the P/D process is the source of the variable frequency spectrum as a function of the level of the input stimuli--not a filter in the signal path associated with a floating model.
+ Excessive Extrapolations of the findings of Hodgkin and Huxley.
Many attempts have been made to explain the functioning of the neuron based on the irritability of a single cell element, the cell wall of the axon portion of the overall cell. This approach is clearly an example of using a "floating model" since the investigators point out the necessity of carefully removing all of the dendritic material associated with the same neuron to avoid experimental instability. If one expands the model to include not only the dendrites of the same cell but the Nodes of Ranvier and other features as well, it becomes apparent that the wall of the axon is a passive structure. It is the junction between the dendrite(s) and the axon that is the active device, designated the Activa in this work, within the neuron.
+ Linearity applied to the general signaling process in animal vision.
The fundamental works of vision in the literature recognize the exponential nature of vision. When one enters the realm of photometry and colorimetry, the community has hypothesized linearity. In the past, hypothesizing linearity greatly simplified the arithmetic and this axiom was formalized as Abney's Law. However, it has been recognized throughout the literature that Abney's Law was not a precise relationship; Abney's Law has a corollary, Abney's Effect, that recognizes the failure of the basic law--and even the corollary as usually stated is known to be in error. This work employs the concepts of the large signal case and the small signal case, with the large signal case being the general case. This work ignores the concept of linearity in the large signal case, reserving it for the small signal case where the mathematical approximation of linearity can always be used if the signal amplitude range is suitably small.
+ The use of the terminology R, G, & B to designate the principle absorption spectra of vision, whether literally or figuratively speaking.
This terminology must be avoided in favor of (UV-), S-, M-, and L- to avoid 'coloring' the discussion erroneously. No "red," "green," or "blue" chromophore exists in animal vision; the actual chromophores are in the ultraviolet (UV-) with a peak absorption near 342 nm. , in the purple-blue (S-) with a peak absorption near 437 nm. , in the yellow-green (M-) with a peak near 532 nm. , and in the orange-red (L-) with a peak near 625 nm.
+ Opponent Theory in its original and most recent manifestations calling for two opponent axes related to the chromatic domain.
This theory based on psychophysical tests, and sometimes only behavioral tests, has consistently attempted to define two essentially orthogonal chromatic axes; nominally red-green and blue-yellow without benefit of a functional model of the visual process. The Hering School generally defines a green photoreceptor to the short side of 532 nm. and a yellow photoreceptor to the long side of 532 nm. Using a comprehensive model, this work will demonstrate the following. The use of the common English names is inadequate. As indicated immediately above, human vision relies on three chromophoric signaling channels that do not conform to these simple and common names; the short wavelength chromophore has a peak absorption in the distinctly purple-blue area at 437 nm. , the middle wavelength chromophore has a peak near 532 nm. and is distinctly yellow-green to the average English speaker. The long wavelength chromophore has a peak near 625 nm. which appears distinctly orangish-red to the English-speaking human observer.
The most useful chromaticity diagram for research purposes, based on this model, will be shown to involve an S- minus M- axis and an L- minus M- axis with a common point defined at the peak absorption wavelength of the M-channel chromophore, 532 nm. This configuration can be extended to include tetrachromatic animals by incorporating a UV- minus S- axis sharing the S- node at 437 nm. This diagram has the unique ability to rationalize the Young-Helmholtz and Hering proposals related to color vision and provides a theoretical foundation for the Munsell Color Space.
For those readers who feel they must continue to embrace some or all of the above hypotheses not used in this work, they are free to do so; however, it behooves them to explain how their alternative can 1) be incorporated into a comprehensive and consistent overall model and 2) provide accurate and precise answers to the performance of the animal visual process over its entire dynamic range.
The biological community has found it difficult to accept the fact that signaling over the neurological system of animals is based on electronic currents. The so-called "gap junction" does not appear as an accepted feature in the neurological literature until the 1960's and later, and then only in a very limited role. These readers may find the presentation that follows difficult to accept, especially since much of the symbology of semiconductor physics will be used to explain the operation of the neuron and the neurological system. Whereas the symbology used here is designed to represent the performance of the actual neurons and not an equivalent circuit or analog, some readers may find it easier to accept the crutch that these are only "equivalent circuits." In this way, they are free to offer other explanations of precisely how the signaling is performed in and between neurons. Clearly, their alternate explanations should indicate exactly how the various voltages and currents measured in the laboratory are generated by purely chemical or biochemical processes.
A few words about Biochemistry versus Bioelectronics are appropriate. Whereas most of the investigators working in biology sprang from the chemical sciences and logically use and rely on a biochemical perspective; the fields of modern electronics and particularly of semiconductor physics provide many tools that are not available in the biochemical field. These tools provide detailed explanations of the spectral, amplitude, power consumption and computational properties of the vision process that cannot be provided through biochemistry. Similarly, the importance of the liquid crystalline state of matter in the operation of the animal organism has not been fully appreciated in the biochemical field. The liquid crystalline state introduces a new realm of physics, electronics and (yes) chemistry to the field of vision research. By introducing these tools, the overall technical field upon which the vision process is based and most completely explained might well be called the Bio-electro-chemical field or bioelectrochemistry.
This work will take the position that the vision system in animals, both sensing and signaling, is based fundamentally on bioelectronics and the role of biochemistry plays a support role. This work will provide a more complete understanding of the vision system and process by showing how bioelectronics, the movement of electrons through the neural structures of the body in the presence of electrical fields, explains all of the relevant activities of vision and that the electrical fields within these neural structures are provided by biochemistry. Other aspects of biochemistry obviously play other, non signaling, roles involved in growth, homeostasis, etc. Using this perspective, most of the processes of vision can be explained using much simpler and more conservative concepts, schematics, and flow diagrams than are found in the current literature.
In the development and concurrent functional analysis of the overall model presented in this work, a necessarily iterative process, several new discoveries have been made, and in some cases even patented.
This iteration of the model has allowed for the continual expansion of the model to incorporate the experimental data found in the literature. This continual iteration and expansion have emphasized the absolute necessity of expanding the fundamental tools and concepts used by the vision researcher and applicable to the model. The expansion has frequently employed subdivision into a very broad general situation and a more limited special situation. Some of these are summarized below:
+ In morphology and cytology, investigators must be very cautious when using the light microscope at or near its diffraction limit to avoid seeing things that are not there; a long standing and well-recognized situation in photo-reconnaissance. It appears (2) that many workers have stained the cells associated with the disk stack of the Outer Segment to obtain a high contrast image under the microscope and then observed the first Airy ring surrounding the disk stack and associated it with a real feature of the cells, specifically a nonexistent cell wall around the disks. Sometimes, the contrast of this Airy ring has been augmented by the presence of other non-membrane materials clinging to the surface of the stack. To understand the actual structure and processes of a cell requires the use of the electron-microscope at x100,000 or higher magnification. Only at this level and higher can the details of both internal and external membranes be identified and understood.
+ Also in morphology and cytology, the juxtaposition of cells is as important as their morphology; particularly with regard to the physical spacing between dendritic and axonal tissue. More specifically, what a cytologist does not see can be as important as what he does see; anytime a dendritic membrane approaches an axonal membrane within 75-100 nm.., it should be carefully noted. In all likelihood, not only is an electrical junction formed in this situation but the junction constituents an active semiconductor device. This is true whether the junction is external to a cell or it is internal to a cell.
+ Every morphological neuron contains at least one active electrolytic semiconductor device, designated an Activa in this work for reasons to be explained later. Many neurons contain several Activas performing similar or different functions. These Activas exhibit nearly identical characteristics to more widely known solid state semiconductor devices, only they were invented much earlier by a higher power.
+ In the neurological system, it is very important to differentiate between the compounds and structures present for metabolism and growth and those present for signaling. The Activa requires the presence of compounds that can supply constant electrical potentials and current, i. e. to operate as batteries. Many calcium-based compounds appear to provide this source of electrical energy, although they do not participate directly in the signaling process. The signaling process appears to be the province of the Activa and the electrolytic conductivity of the materials inside a given cell wall.
+ The electrolytic foundation for the electrical circuits of the neurons introduces two extremely important realizations: the combination of a rechargeable battery and a diode in series provides an entirely reversible impedance that can be used in an amplifier circuit. The resulting circuit does not contain any dissipative impedance elements (except parasitic elements) and operates as a completely reversible thermodynamic process; no power is consumed and no heat is liberated. This is the answer to the remarkable thermal efficiency of the neurological system.
.The conductors of the neurological system consist of electrolytic channels instead of metallic conductors, with signal propagation measured in 1-10 meters/sec instead of 108 meters/sec as in metallic paths. The finite signal delays associated with the various signal paths must be accounted for when attempting to introduce lumped constant filter theory into the explanation of the operation of the eye.
+ In interpreting the signals involved in animal vision, expanding the concept of signaling into the two cases, the large signal case and the small signal case is very important. As in any mathematical formulation of a nonlinear process, the fundamental large signal case can always be limited sufficiently to allow a linear formulation of the derivative small signal case. Recognition of this situation allows one to rationalize the obviously logarithmic relationships of the large signal case, Beer's, Fechner's, Weber's, etc. Law with the attempts to describe vision as a linear process in the small signal case; Abney's, Grassman's, etc. Law.
+ This author has been continually surprised at the limited application of the overall concept of feedback in the vision literature; especially the preponderance of attention given to external feedback to the apparently complete exclusion of the more frequently intrinsic, and more easily implemented, concept of internal feedback. Similarly, the vision literature seems to concentrate on the less specific biological/psychophysical concept of inhibition where the broader concept of subtraction leads to a much more comprehensive description of the resultant signals.
+ The literature of vision, even in highly specialized presentations, has ignored the general equations of optics in favor of the Gaussian Approximation. This has caused the publication of hundreds of incorrect figures showing off-axis optical rays passing through the nodal point and additional figures implying the image on the retina is a linear copy of the object. The Gaussian Approximation is accurate enough for commerce, specifically optometry; it is not accurate enough for scientific research in vision. Failure to use the general equations when necessary has deprived the community of many valuable insights into the optical system of the eye; i. e., the variable focal length of the optical system, the high degree of spatial non-linearity in the image projected onto the retina, and the importance of the variable index of refraction incorporated into the lens of the eye has not been appreciated. Of even greater significance in evolution is the fundamental difference in operation of the optical system of marine animals and terrestrial animals and also the specializations employed by animals that live in a niche at the interface of these two environments.
+ It is frequently lost in the discussion but important to recognize that the work of the C.I.E. and its predecessors was not aimed at or to support research; the primary goal of the Committee was to provide photometric and chromatic tools for use in commerce and engineering. The C.I.E. provided a relatively simple, highly filtered by smoothing, and eminently useful small signal interpretation of the visual process. The purpose of this work is to provide a more exact, unfiltered and more detailed large signal interpretation of the visual process for use in research.
+ Whereas at the time that it was ordained that a derivative of Vitamin A conjugated with the protein opsin via a Schiff base to form the chromophores of vision, there were only a handful of derivatives of Vitamin A known and the number of ways two ligands could combine could be counted on the fingers of one hand. Now, more than a thousand retinoids are known, many of which can be considered as derivable organically from Vitamin A, and entirely new methods of bonding are recognized. This has allowed the designation of a specific homologous family of resonant retinoids, designated the Rhodonines, as the chromophores of vision when in the liquid crystalline state. Members of this family are derivatives of Vitamin A. Their association with a particular protein substrate can be shown to be via hydrogen bonding and much less important in the visual process than thought previously.
The vision community has completely overlooked the importance of differences in time within the visual system. This is understandable based on its heavy emphasis on morphology to determine the topography of the visual system. Using morphology, it has been able to determine the role of the LGN, basically through what might be called "traffic analysis" in the intelligence field. However, it has not been able to explain how the LGN functions. By introducing the concept of differential time delay, both with respect to the electrons within the disks of the Outer Segments (to provide the solution to the Photoexcitation/De-excitation equation) and between signals in related signal paths between the retina and destinations within the brain, a much more complete understanding of the visual system results.
Zeki has addressed a serious problem in the vision literature associated with language and semantics. Lacking a strong central secretariat, each author has taken considerable license with the (generally) English language. Short expressions are frequently used to describe complex relationships for convenience. When these short expressions are referenced by later authors, the short expressions are frequently shortened further or interpreted differently than the understanding of the original author.
This author is sometimes accused of using "old data" from the literature. It has frequently been necessary to return to original sources to determine what the un-extrapolated measured facts are. Two important examples are worth mentioning at this point:It is also important to review original documents to discover what physical foundation was used in the work. When exploring neurology, it is important to note that Nernst was not dealing with biological membranes. Neither was Goldman. He manufactured non-biologically viable membranes from previously viable materials. Hodgkin, Huxley & Katz noted almost immediately the negative impedance that was measured in their early experiments; a sure sign that there was a battery buried in the electrical circuit of the specimen--and more importantly-- that Ohm's Law did not apply to these measurements. The more fundamental basis for Ohm's Law, Kirchoff's Law should have been applied.
It is also important to address the subject of (Annual) Review Articles. Most review articles are designed to provide an overview of recent activity in a given field. They are a valuable source of references. However, they invariably provide superficial interpretations of the work of a large group. For the sake of brevity, they frequently omit a variety of important adjectives and constraints relative to the data. They normally include an introductory section relating to the generally accepted wisdom. It is seldom supported by references. They are frequently prepared, by request, by an author with a parochial perspective and support his agenda.
In cases where an investigator or an author of a text,
it has been necessary for this author to reference original sources to reestablish the factual trail. Other investigators are encouraged to follow this necessarily laborious approach.
The author has more than 30 years experience as a system engineer in imaging and scanning systems. The primary focus has been on the synthesis of high performing imaging systems for reconnaissance.
His early work involved creating electronic imaging cameras that could outperform photographic film cameras. This work was the first to show that the resolution of film was defined by the distribution of grain size as described by a log-normal, not Gaussian, distribution. It also determined the theoretical quantum efficiency of silver-halide films, and that granularity was in fact quantum noise in a nominally 1% efficient process.
During the early development of night-vision goggles, he showed that the quantum efficiency of the goggles was actually poorer than the human eye; the advantage of the goggles was in their optical gain (due to aperture size) and electronic gain which together made their output image more compatible with the state of the soldier's level of adaptation under field conditions.
It is this author's opinion that progress in the visual science field has been very slow compared with other sciences for three principal reasons:
Progress has probably also been slowed by the normal homocentric viewpoint of many senior investigators who have assumed and supported the viewpoint that human vision is the most effective in the animal kingdom.
In the Preface to volume VII/6B of the Handbook of Sensory Physiology, Autrum opens with the statement that:
Even with his amplification, the thought is still too limited. A more comprehensive phraseology would be:
This restatement of Autrum's quotation has two objectives illustrated in Figure P-1; first, to show that morphology and physiology cannot stand alone as a pair in the exploration of the vision process, they must actively interface with a dditional fields, particularly bioelectronics. Second, they must interface with the appropriate analytical tools of these other fields. In the above context, morphology is clearly a euphemism for the fields of anatomy, morphology, histology, and cytology.
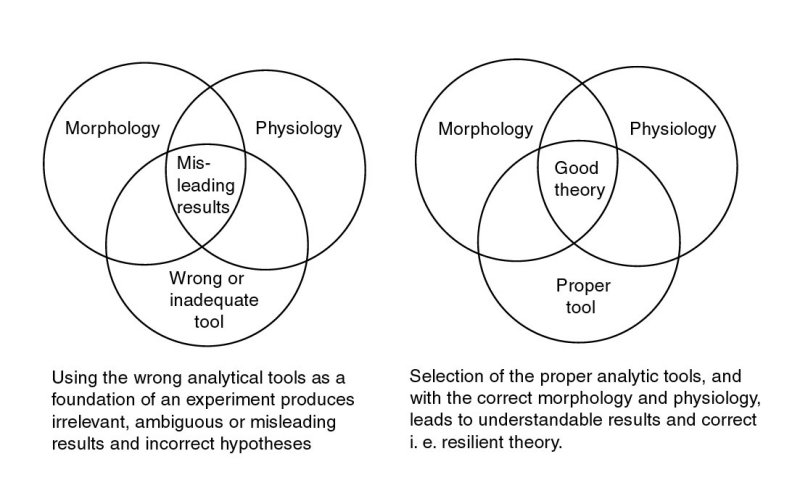
Although many crucial examples exist in vision research where the investigator has failed to use the appropriate analytical tools, only one illuminating example will be cited here.
Recently a highly competent research team carefully designed and carried out an experiment recognizing the correct morphological and physiological conditions but made an error in their selection of analytical tools. They assumed that the operation of the retina depended on the dissipative character of the laws of irrereversible thermodynamics when in fact the retina is based on the completely reversible thermodynamics of electrochemistry. Their experiments led to negative results; which was both fortunate and intellectually illuminating. It was fortunate in it did not lead to new misleading hypotheses. It was intellectually illuminating in that it showed that no significant dissipative elements (resistors) exist in the retina to generate heat. The team went on to redesign their experiment and look for concentrations of electrochemical activity instead of heat.
This work is the result of the interest of the author in the dynamics of vision, not merely the static characteristics. No major document could be found that discussed the dynamics in a comprehensive manner. Nearly all of the larger texts in the field are in fact compilations of facts grouped together in chapters. These chapters lack a continuity, a defining thread, a consistent end to end model. They also tend to avoid the dynamic aspects of vision. The result is a low level of understanding of vision. Figure P-2 presents a chart of facts versus understanding, using semi-logarithmic coordinates, that may be helpful. The assumption is that the horizontal axis represents understanding and understanding requires analysis. The vertical axis is related to the total number of facts known in a given field. The curve seen expanding upward to the right is the level of knowledge gleaned from the total milieu of facts at a given time and incorporated into a framework of understanding. In this context, exploratory research increases the total number of facts known in a given area; and in other areas as well. This research leads to an increase in knowledge. It does not lead to an increase in understanding. Understanding is the result of employing research to confirm a hypothesis based on analysis. This leads to a consolidation of knowledge in a framework of understanding.
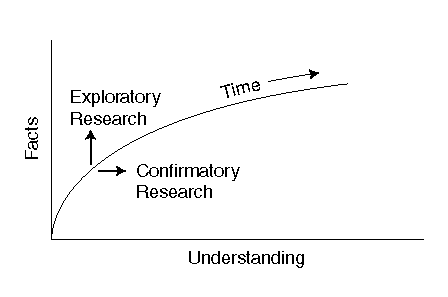
This figure conforms roughly to the normal mode of teaching. Proceed first in the vertical direction to provide knowledge (who, what & where). When the student is ready, proceed in the horizontal direction to provide understanding (when & how). It is hoped that this work provides a greater measure of understanding.
After reviewing a variety of books on neurology and neuroanatomy, an interesting conclusion can be drawn. They all profess to discuss the morphology and the function of the neural system. However, the portion on function is invariably limited to what the intelligence field (CIA, etc.) would define as traffic analysis, the description of the number of neurons connecting various points and some discussion of the speed of signaling over those neural signaling channels. Although there is some discussion of the polarity and potential of the various plasmas (primarily the axoplasm), there is virtually nothing on the actual processes and mechanisms associated with the neuron. What material claims to portray the underlying mechanism of the neuron is based on very elementary and inadequate physics and chemistry. Marr makes the key observation that neurophysiology and psychophysics have as their business to describe the behavior of cells or of subjects but not to explain such behavior. While the term function frequently appears in titles for marketing purposes, it should normally be replaced by the term taxonomy, as used in Uttal's book.
Morphology has occasionally failed the biological investigator; a shining example is assigning the name amacrine, meaning anaxonal or without axon, to certain cells in the retina. Morphology itself, depending as it does on essentially exterior dimensions, could not discern the fact that the dendritic and axonal structures were aligned side by side in essentially one cylindrical structure. This structure could be more appropriately labeled the dendrite/axon "process." This structure ("process") is known through physiology to both accept and deliver signals to other nearby cells.
In this age of specialization among scientists, the average scientist (even the leaders in a field) read certain journals, attends certain conferences and meets with the same groups of specialists year in and year out. In this atmosphere, orthodoxy matures and cross-fertilization is suppressed. The same individuals who formulate the orthodox theory enforce it by reviewing papers submitted to the journals and grant proposals. Paraphrasing Nobel Laureate Hannes Alfven, Galileo was not so much a victim of the Catholic Church as he was a victim of peer review. Kuhn has discussed this question in detail
We are all aware of the game of sending an article to a journal controlled by a "peer" favorable to our ideas. Unfortunately, the broader, the paper, the more difficult to find a journal with an agreeable peer. Quoting Gutmann & Lyons introduction in 1981, "Chemists are often not familiar with the conventional solid state concepts...." None of them are familiar with the even more sophisticated concepts of electrolytic semiconductor theory.
The system of peer review--having all papers and grant proposals controlled by a small group of "leading specialists"--has had a profoundly conservative effect on all branches of science. The ultimate test of scientific theories is observation. I will emphasize how observations conflict with, or support, various earlier ideas. I welcome experiments to confirm, disprove or better characterize the ideas proposed in this work. The Appendix contains an extensive list of suggested experiments. Some experiments are entirely new, many repeat earlier work under more rigorously controlled conditions.
Anyone who conducts an argument by appealing to Authority is not using his intelligence, he is just using his memory.
Leonardo de Vinci (1452-1519)The virtual, if not complete, absence of the discipline of solid state physics (use the expression solid state chemistry if preferred) from the literature of vision is startling. A researcher with a broad background would find this situation inconceivable after reviewing the clearly semi-crystalline (more properly liquid crystalline) nature of the disks of the photoreceptor cell Outer Segments--as well as most other structures in the eye. This situation is almost certainly due to the limited technical breadth of the "peers" controlling the direction and aiding the planning of research in the field of Vision.
Uttal explored this problem in the above work (page xix) where he decided to re-define the pejorative title "speculative psychologist," bestowed upon him by the vision community, into the honorable task of accumulating the available data, elucidating the possible underlying mechanisms and then putting it all together into a coherent and consistent explanation. I share his emotional task but prefer to use the title "system electro-physiologist" based on the approach taken in this work.
This work can be considered an alternate and unfounded Theory of Vision written by an "outsider." However, it is the only theory available today that:
As an example, it is the only work that describes (much less attempts to describe) the phenomena of color constancy. It also provides the only known description of the mechanism of adaptation and it provides the first known theoretical derivation of the luminous efficiency function for all four illumination intensity regions. It also demonstrates that the C.I.E. (1951) Scotopic Luminous Efficiency Function actually applies to the mesopic intensity region.
To stress the peer problem, Uttal, who is well known in his community, felt it necessary to "question" (refute) 18 Dogmas he found associated with his field of interest in 1981. He then lists 14 factors contributing to contemporary overcommitment to empiricistic neuroreductionism (page 719-743). Amen!
As Burton pointed out in his introduction to circulation, "Physiologists would be unlikely to agree on details of any statement, but the following would be given general approval: . . ." He submitted his work to a committee of his own creation, the Celestial Committee on Design of a Mammalian Circulation. It is suggested that comprehensive opinions concerning the value of this theory be withheld until after the theory has been tested in the vision community. However, the author will be happy to respond to any written or verbal queries at any time.
Theoreticians and investigators alike are cautioned to discount gratuitous pseudo-authoritarian and sweeping statements made by eminent members of the community. As an example, one writer hinted at the necessity of random interconnections in the INL of the retina "due to the limited amount of genetic code available." Since no one has claimed to know the algorithms involved, it could be a fractal code consisting of a constant and an equation of less than six terms. How someone could justify the above quotation without some numerical description of the amount of the genetic code involved in the vision subsystem of the animal, any animal, is hard to comprehend.
Another problem presenting a dichotomy to this author is the currently raging argument over parallel channels in the visual system. It primarily involves the community studying the cortex. However, one can encounter highly volatile, even vituperative, responses from some quarters if a position is taken on either side of this issue. To be clear, this work presents a thesis that straddles the fence. It has found the presence of highly defined channels in the pre-cortical area necessary and defendable. On the other hand, it has found virtually no defined parallel channels in the cortical area. It has gone further and pointed out the lack of sufficient knowledge to even address the question in the cortical area.
On the other hand, this author acknowledges the meticulous laboratory work of so many investigators who have contributed unwittingly (and over many years) to the development and corroboration of this model.
Laboratory techniques of the 1930-50's were a major step forward from earlier times; even with the tools of that day, understanding expanded greatly. Many axioms (hypotheses) put forward then have been accepted for 40 years. Many new tools have become available meanwhile, particularly through interdisciplinary approaches in fields like the photochemistry of photography, the circuit theory of electronics and the solid state, the discovery and development of the liquid crystalline state and even the expansion of the field of chemical bonding to include the hydrogen bond ( previously Van der Waal bonding).
These new analytic tools and the associated advances in laboratory technique, such as nuclear magnetic resonance, are now able to provide a much more extensive and internally consistent theory of animal vision than was possible earlier. That is the purpose of this work.
To close this section, it is useful to consider the philosophy of my former friend and colleague, and a great teacher, Professor Dennis Gabor of Imperial College. I subscribe to it.
In the course of formalizing this work, including documenting the fundamental tetrachromatic nature of animal vision, several fundamental discoveries have been made:
I hope the current investigators will find the model presented herein a framework for explaining their work and a useful point of departure for future work. The author has received great pleasure from the puzzle-solving accomplished to date. However, the further elucidation of the work, mopping-up in the language of Kuhn, will require many hundreds of man years. I must enlist the aid of a broader community to complete this task. Much remains to be done.
| I am never content until I have constructed a (mechanical) model of what | |
| I am studying. If I succeed in making one, I understand; otherwise, I do not. | |
| William Thomson (Lord Kelvin) |
Return to the website home page
1. Jeans, J. The Growth of Physical Science. pg. 237
2. "It appears" is a signal to the reader that the following statement is based on logic and is not, as far as the author knows, substantiated by careful experimental evidence.
3. Autrum, H. (1981) Comparative Physiology and Evolution of vision in invertebrates--B; Invertebrate visual centers and behavior I. NY: Springer-Verlag
4. Uttal, W. (1981) A taxonomy of visual processes. Hillsdale, NJ: Lawrence Erlbaum Associates.
4. Kuhn, T. (1972) The structure of scientific revolutions, 2nd ed. IL: University of Chicago Press, Chapter 3