
This page is in beta release. The author welcomes and will respond to any comments or suggestions left at the comment page. Section numbers of the main manuscript, available on the web, are shown in brackets. The manuscript can provide more detail when desired. The first number shown is the chapter number; it is followed by the section numbers. Download individual chapters.
Adaptation is a term used to describe the remarkable ability of the visual system to automatically adjust its sensitivity to accommodate a range of illumination greater than 10 orders of magnitude(efficiently over more that seven orders). This adaptation is achieved largely without the persons knowledge. To achieve this operating range, several distinct mechanisms are involved. Each of these mechanisms exhibit separate time constants that can be demonstrated under different experimental conditions. These time constants cover a very large range, from two milliseconds to ten minutes and appear differently during light adaptation (adjustment to accommodate increases in light intensity) and dark adaptation. Their significance in a given situation is also a function of the amplitude of the change in light intensity. The spatial acuity and color performance of the visual system are closely linked to the level of adaptation but are generally associated with different underlying mechanisms.
The following material is divided into sections:
The phenomenon of adaptation is complex and involves many individual mechanisms. All of these mechanisms are associated with the photoreceptor sensory cells of the retina. The individual mechanism involve time constants that vary over many orders of magnitude and come into play individually at different levels of retinal stimulation.
When adaptation was first conceptually recognized is lost in history. Behaviorally, it must have been recognized by the earliest people. Significant effort was expended by Hecht, et. al. in the 1920's and 30's to quantify adaptation using psychophysical methods. However, the instrumentation of that day was limiting in many respects. More recently, electrophysiological methods have allowed more extensive investigation of the phenomenon, particularly at very short time intervals after a change in the stimulus level. Because of ethical considerations, most of the best data has come from research on other high level primates whose visual systems are functionally very similar to that of humans.
Because of the complexity of the overall phenomenon and the complexity of the mechanisms creating the phenomenon, it is difficult to correlate all of the findings of the psychophysical literature, the electrophysical literature and the physiology of the visual system. Only recently has the physiology of the system been defined in sufficient detail to allow a comprehensive description of the overall phenomenon.
The overall phenomenon exhibits different characteristics depending on whether the light stimulation level is in the mesotopic, photopic or hypertopic regimes.
Adaptation is best investigated when the underlying physiology is modeled as shown in Figure 1. The figure shows light passing through the pupil of the eye as determined by the iris. The light excites the chromophoric material on the disks of the outer segment of one of the photoreceptor cells. The energy associated with the excitons generated in this process is transferred to the dendrites of the first amplifier circuit within the sensory neuron. This circuit is labeled the adaptation amplifier. The energy transferred to the the dendrites participates in an amplification process very similar to that found in man-made transistor circuits. The process generates free electrons that are transferred to the next circuit within the neuron via the dark lead at lower right. The circuitry shown at upper right represents the power supply supporting the adaptation amplifier. This circuitry plays a critical role in the adaptation process.
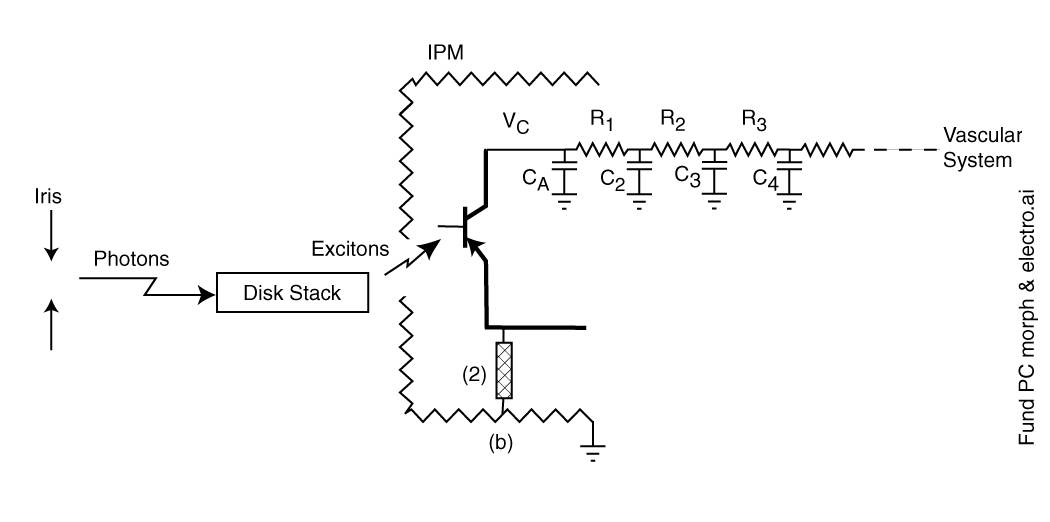
Figure 1 Elements participating in the temporally dynamic aspects of vision--adaptation
Adaptation begins with the control of the light entering the eye by the iris. While the main purpose of the iris is to control the spatial quality of the image formed by the lens on the retina, its opening and closing does provide a crude adaptation mechanism over a range of only 4:1 to about 16:1 on a time scale of seconds [2.4.3]
The disk stack is the structural element known physiologically as the outer segment of each photoreceptor neuron. This element consists of about 2000 individual disks of the structural protein known as rhodopsin. Each disk is about two microns in diameter and 200 Angstom thick on average [4.3.5]. Each disk is coated on all sides by one of the chromophores of vision deposited in liquid crystalline form. These disks are physically isolated from each other for quantum-mechanical reasons [4.3.5] but are in common contact with one or more dendrites associated with the first amplifier in a single photoreceptor cell. This amplifier is built around an electrolytic semiconductor device known as an Activa.
. . .The sensory neuron is shown using conventional transistor device symbology along with the electrical analog form of its support elements.
The impedance level of the electrolytic photoreceptor neuron is orders of magnitude higher than any electronic circuitry made by man. As a result it is very difficult to measure the voltages associated with these circuits with conventional electronic instrumentation.
Elements R1 and CA are conventional electrical elements related to the capacitance and resistance associated with the plasma wall of the sensory neuron. They are reported to provide a time constant of about 0.1 seconds to the overall circuit.
Elements R2 and C2 represent the hydraulic impedance of the neural substrate of the retina. These elements together exhibit a time constant of about two minutes. Elements R3 and C3 represent the hydraulic impedance of the optic artery and veins. They together exhibit a time constant of about ten minutes. C4 represents the large capacity of the vascular system to support vision. This capacity is so large that no time constant can be assigned to it for the present purpose.
These elements exhibit a series of time constants that only come into play when the actual or equivalent voltages at their respective nodes deviate from their equilibrium value substantially. Thus, the overall adaptation characteristic of vision varies significantly depending on the level of light stimulus introduced (over a range of at least three and one-half orders of magnitude. Adaptation occurs both in response to the application of light and following the termination of that light. Adaptation plays a key role in color constancy when a broad spectral band light is used as the stimulus. However that subject is addressed elsewhere on this site.
The adaptation process can be broken into two major processes,
Dark adaptation is much more frequently discussed but light adaptation is of equal if not greater importance. Light adaptation is an active defense mechanism designed to prevent damage to the highly sensitive visual system by excessive light levels. Dark adaptation is a return to high sensitivity operation in the absence of an excessive light level.
Light adaptation is accomplished in an active mode by three different mechanisms.
. . . . .The operation of the overall adaptation phenomenon can be described by describing the role of each of the elements in the above figure.
Figure 2 shows the transient performance of the fundamental Photoexcitation/De-Excitation (P/D) mechanism in vision. This characteristic is exhibited by every photoreceptor. It is associated with the excitation and de-excitation of the electrons within the liquid-crystalline chromophore coating on the disks.
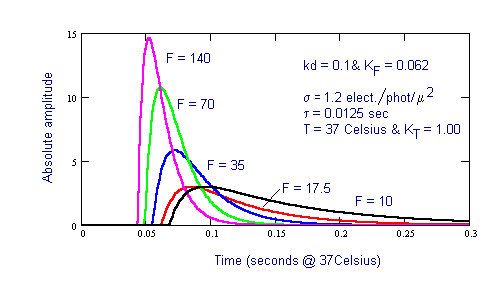
Figure 2 Fast adaptation process associated with the Photoexcitation/De-excitation process in the chromophores
The intensity, F=10, in the above figure corresponds to the lower edge of the photopic region of vision. Note the significant change in the slope of the on-transient response with light intensity, F. The off-transient or decay transient shows much less change in slope with light intensity. Note also the significant delay in the response of the chromophores to light depending on the intensity of the illumination. For the mesotopic and scotopic light regimes, the on-transient response has a fixed time constant of 12.5 milliseconds. The on-transient time constant falls with light level throughout the photopic region, eventually reaching a value below 1.0 milliseconds. The off-transient of the P/D mechanism is 12.5 milliseconds at all light levels above the mesotopic/photopic transition but becomes longer in the mesotopic and scotopic operating regimes.
The impedance of the adaptation amplifier circuit within the sensory neuron plays a significant role in adaptation whenever the light intensity is sufficient to cause a shift in the DC value of the axon of the neuron. This circuit has a time constant of about 0.1 seconds. This time constant is derived from frequency response data for the human eye. It exhibits a easily recognized frequency pole at three Hertz which can be equated to a transient time constant near 0.1 seconds.Figure 3 shows the temporal response of the iris. The time constants involved are of the intermediate class in the overall description of adaptation.
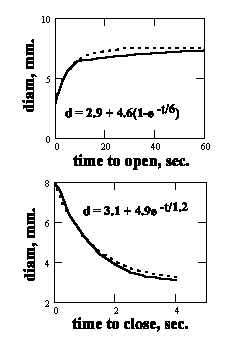
Figure 3 The transient performance of the human iris
The primary role of the iris is maintaining optimum spatial quality in the image projected onto the retina by controlling the "speed", f/#, of the optical system. Its adaptation range is quite limited, between 4:1 and 16:1 depending on the investigator. This limited range is generally disregarded in discussions of adaptation. [2.4.3]
Figure 4 illustrates the transition of the slow adaptation process associated with the vascular system from a simple exponential function to an exposine function. Following exposure to light under photopic conditions, the visual system will recover following the exponential curve shown. Following exposure to light under hypertopic conditions, the system will exhibit the more more complex exposine response shown.
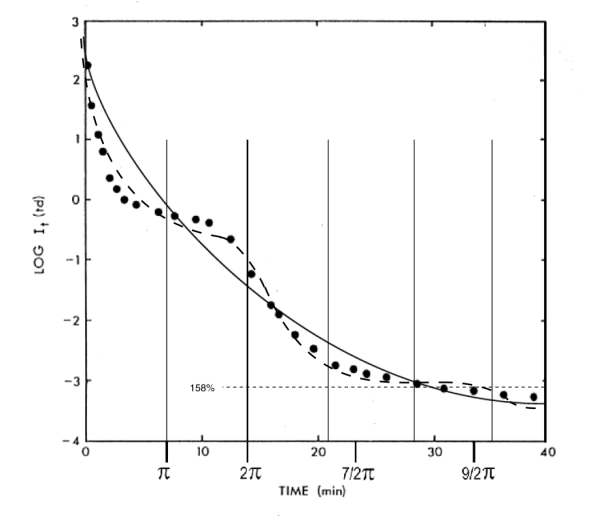
Figure 4 Exponential to exposine function transition in adaptation mechanism associated with the vascular system
More extensive correlation between the theoretical model of adaptation and measured performance is shown in Figure 5.
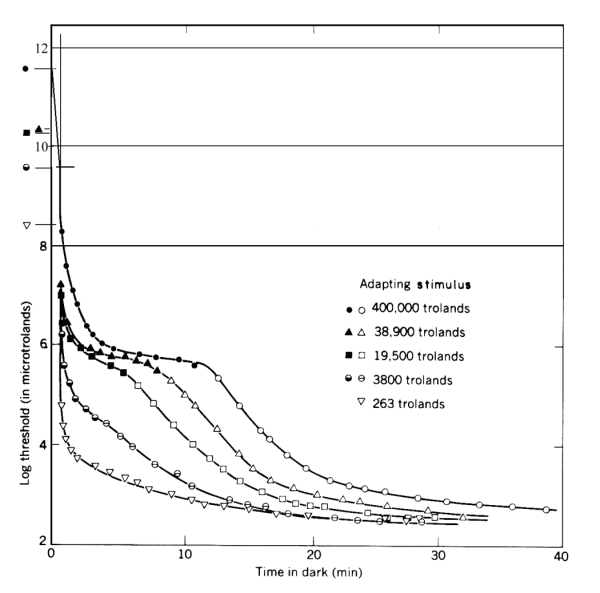
Figure 5 Comparison of the theoretical performance and the measured data base from Hecht, et. al.
This figure may appear unusual to some. It normalizes all of the responses to a common ultimate sensitivity. As a result, the starting point of each transient is at a different level along the ordinate. The resulting presentation shows the change in illumination level associated with each response.
The middle curve in the above figure is near the upper extreme of the photopic region. The upper two curves are only obtained under hypertopic conditions. "Snow-blindness" and "beach blindness" are typical situations of hypertopic adaptation. Full recovery (full dark adaptation) following encountering these conditions for only a few minutes can require several hours.
1
2