
The fundamental morphology/cytology of the neuron has not been well represented in the literature. This is primarily due to the great variety in neural forms and their very high aspect ratio. There are a profusion of caricatures at the conceptual level but little precise information concerning the relevant detailed cytology of the cell.
This page is divided into three sections:
The external morphology of a typical neuron
The internal circuitry of a typical neuron
The cytology of the fully elaborated neuron
To understand the cytology and operation of the neuron in detail, refer to the text:
Biological Vision: A 21st Century Tutorial, $25.99 USD. Ships from stock.
The figure below illustrates a simple pyramid neuron grown in culture. It shows the axon (arrow) extending to the left and several neurites converging on the neuron. As will be discussed below, it is proposed that the top two neurites are dendrites (D), the smaller one on the left, and the lower neurite is a podite (P).

The primary fact is that every neuron contains at least one Activa. This Activa is an active analog electrolytic semi-conducting device. When combined with the appropriate circuit elements, it can be made to perform a variety of functions, including but not limited to oscillation.
The following figure shows a minimal neuron [10.6.1]. It will be defined as the nominal or FUNDAMENTAL NEURON for purposes of this work. The cutout in the lower left shows the symbolic representation of the Activa overlayed on this same neuron.

The figure is shown with two distinctly separate dendritic roots. This is unusual. It is also shown with one poditic root and one axon. In this configuration, the neuron is frequently described as star shaped (more than three limbs). The truly fundamental form is frequently labeled a pyramid cell (one dendrite, one podite and one axon). All other forms are repackaging of this fundamental form for secondary reasons such as available space, convenience or the requirements associated with auxiliary components.
All neurons incorporate Activas and Activas are three terminal electrolytic devices. In signaling analyses, it is customary to separate the electrical paths between these three terminals and the surrounding common electrical "ground" into two separate groups, those paths that carry the signal information and those paths associated with providing electrical bias voltages to the device. Sometimes the paths are shared but it is always useful to make this distinction. In the following discussion, only the signal terminals on the surface of the neuron are addressed.
Although of equal importance to the signal terminals, the bias terminals of neurons are essentially unrecognized in the literature. They are addressed in depth in the main text. They are intimately involved with the electrostenolytic process that powers all neurons via the glutamate cycle of metabolism.
In the drawing, the terminals marked A through D are all positive inputs at synapses. They are defined as positive in the sense that the output signal associated with the axon is of the same polarity as the input applied at one of these locations. This output is an analog waveform in the absence of other components. Signals applied to terminals marked E and F are negative inputs. The output signal at the axon will be of the opposite polarity compared to these signals. These letter designations are located similarly to a ett used by Guyton where he described locations A through D as excitatory and E and F as inhibitory. These are poor terms to use when speaking of the fundamental analog neuron since they imply an oscillatory or switching function.
The fundamental neuron is a very small morphological structure relative to its electrical capability. In its basic form and size, the device will not oscillate or switch its state of output. However, there are a number of conditions that can make this circuit become unstable. Two of these conditions are critical. If there is a finite impedance in the circuit between the base terminal of the Activa and the surrounding medium, negative internal feedback is introduced into the circuit. This alone will not cause the circuit to become unstable. If a reactive impedance, namely a capacitance is introduced between the collector terminal (less frequently the dendritic terminal) and the surrounding medium, the combination of the capacitance and the negative internal feedback can lead to instability.
Capacitance is normally added to the collector terminal of the above neuron due to the simple need to make the axon longer in order to transmit its output signal to a desired location. As the length of the axon exceeds a fraction of a millimeter (or an experimenter adds capacitance associated with his probe), the circuit becomes unstable. It will either oscillate intermittently or continuously depending on the voltage between the emitter and base terminals. This is the methodology for generating action potentials using an analog device.
Many investigators have attempted to locate the source of the instability within a neuron. Because of the difficulty of measuring the voltage associated with the dendroplasm, the norm has been to trace back from the synapse of the axon. This procedure has usually located the source, frequently labeled the "trigger zone," as near the point where the axon attaches to the cell body. This area is known as the hillock of star and pyramid shaped neurons. It is less well defined in other types of neurons. The Activa of the FUNDAMENTAL NEURON is located at the junction of the dedrite conduit and the axon conduit within the neuron. It is the unusual surface topography of part of the hillock that defines the precise location of the base region of the Activa.
One millimeter is a short distance in a large animal. If the axon is made significantly longer than one millimeter, the capacitance associated with its extremely thin lemma rises quickly. This causes the impedance of the overall circuit to be reduced. The resulting power requirement for achieving a given voltage output rises unacceptably. Beyond a fraction of a millimeter in length, the surface of the axon is usually myelinated in order to minimize the circuit shunt capacitance. This allows the generation of a large voltage action potential using minimum power and to transmit it a significant distance without excessive attenuation.
The attenuation associated with a long axon is not related to the resistive elements associated with the axon. These real resistive elements are negligible. However, any cylindrical electrical structure exhibits both a capacitance and an inductance. These parameters are functions of the dimensions of the structure.
For signals at frequencies above zero, the combination of the inductance and capacitance exhibit two important characteristics. They exhibit an effective resistive impedance at the input to the structure and they define a dispersion characteristic that degrades the signal as it travels along the structure. It is this dispersion factor that the biological investigator has usually mistaken for a resistive attenuation.
The following figure provides a detailed schematic of the completely elaborated neuro-secretory cell, the neuron. The schematic is typical of the sensory neurons, such as the photoreceptor cells of the retina or the hairs found both externally and within the auditory system. It also represents the secretory cells of the intestinal tract.
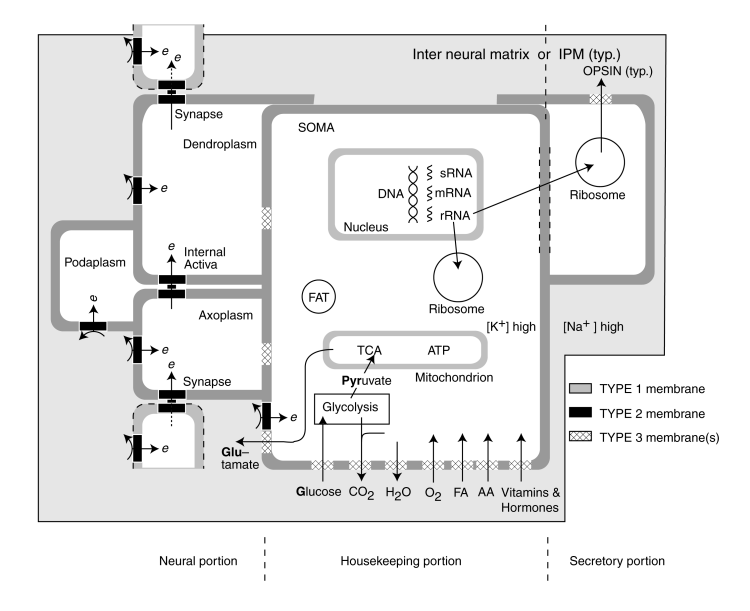
The figure is divided into three portions; the housekeeping portion in the center, the neural signaling portion on the left, and any secretory portion on the right. The relationship between the neural and secretory portions is beyond the scope of this page.
The housekeeping portion is aerobic and involves the absorption of a variety of materials from the surrounding inter neural matrix (INM). These materials are used both to build the cell and maintain the cell.
In addition to its routine aerobic functions, the housekeeping portion performs the anaerobic function of creating glutamic acid (glutamate) to power the neural portion of the neuron. It does this by absorbing glucose and converting it into pyruvate by glycolysis. The pyruvate is processed further within the tricarboxylic acid cycle (TCA and also known as the Kreb cycle or the citric acid cycle) to form a precursor to glutamate, a-ketoglutarate.
An extended version of the TCA cycle, including what is called the glutamate-GABA shunt is used to convert the a-ketoglutarate into glutamate. Glutamate is shown leaving the housekeeping portion of the cell at the lower left. After release into the INM, the glutamate reacts at the electrostenolytic sites (diagramed using the curved arrows). These sites use type 2 biological membranes, BLM's, to decompose glutamate into GABA (gamma amino butyric acid)and carbon dioxide. In the process, the excess energy is used to inject a free electron into the plasma forming the various conduits associated with the signaling function of the neuron. This is the source of the negative electrical bias observed in all mature living cells.
Following its generation, GABA is returned to the TCA by another process within the glutamate-GABA shunt. This process converts GABA into succinic acid, a well known chemical within the TCA, for reuse.
The neural portion consists of three electrically isolated conduits, each with its own plasma biased separately, separated from each other by an internal active region containing an Activa. The dendritic conduit is in active communications with the preceding axoplasm via a synapse (also containing an Activa. Similarly, the axoplasm of the neuron is in electrical communications with a subsequent neuron through its synapse. The previously undocumented third conduit is also a neurite and is designated the podite. This third conduit plays a crucial role in the operation of every neuron.
As noted earlier, the active electrolytic semiconductor devices, the Activa within the neuron and at the junctions between neurons, are used to transfer electrical charge from one conduit of a neuron to the next as shown by the vertical arrows on the left. This movement of charge (resulting in a change in voltage) constitutes the signal transmitted by the neural system.
In the case of sensory or digestive neurons, the secretory portion of the neuron on the right accepts raw mateials from the housekeeping portion and creates one of a variety of proteins. In the case of the photoreceptor cells, the protein is opsin, the basic material used to form the disks of its outer segments.
To understand the cytology and operation of the neuron in detail, refer to the two texts:
Biological Vision: A 21st Century Tutorial, $25.99 USD. Ships from stock.
or
The Neuron and Neural Networks (available in September 2010)
For a broader discussion of the neuron on this website, go to the Neuron Index Page.