This animation is the first of two that develop the method of, and the underlying mechanism supporting, generation of the neural signals from the acoustic energy applied to the oval window of the cochlea.
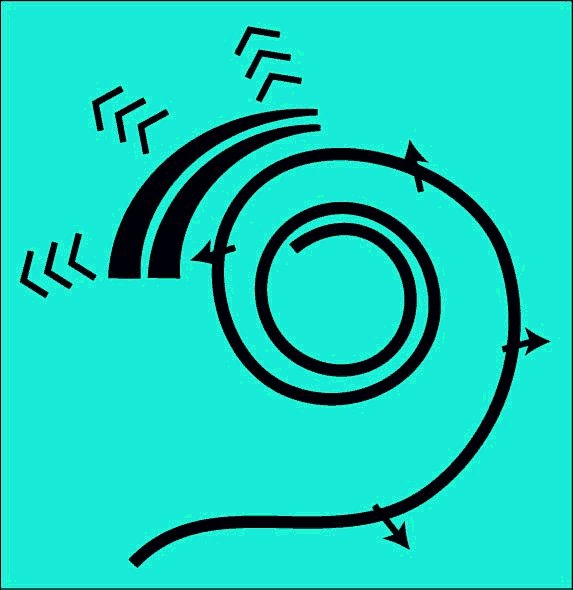
The underlying mechanism requires a curved cochlea. A straight cochlea is not a functional cochlea. While the cochlea need not be coiled as in the higher mammals, it must be curved. It is the local curvature of Hensen's stripe that provides the mechanism for frequency component separation. While birds typically have a cochlea containing less than one-half turn of a spiral helix, the monotremes have less than one and one-half turns, and humans have about two and one-half turns, other animals have up to four turns. The following animation is shown straight for pedagogical purposes only. The next animation will discuss the critical role played by curvature.
The fundamental task of the cochlear partition is to generate two distinct classes of neural signals. One set of signals is based on the tonal information related to individual frequency components of the applied energy. The other set of signals is based on the information describing the envelope of the applied energy. Comprehension of speech and other communications signals cannot be accomplished satisfactorily without the proper operation of both the tonal and the envelope (or temporal) portions of the hearing system.
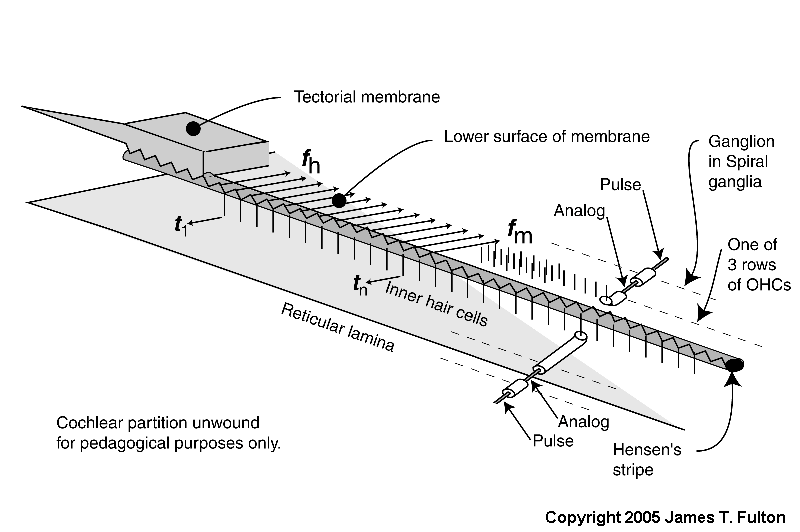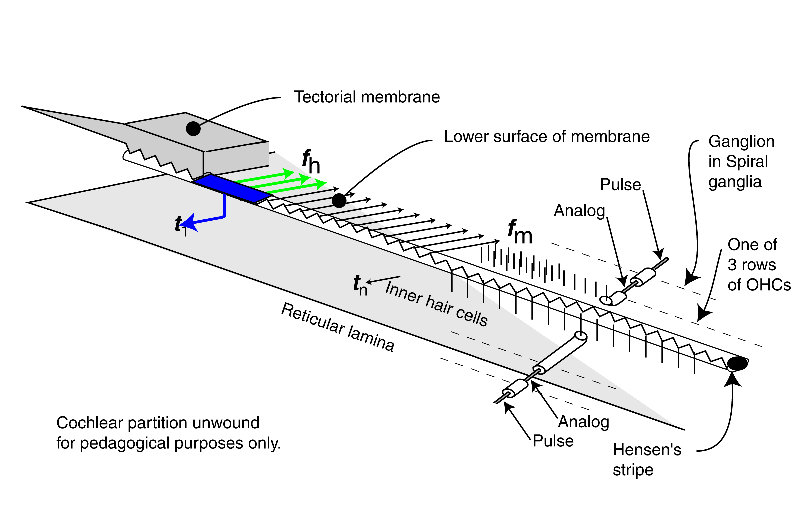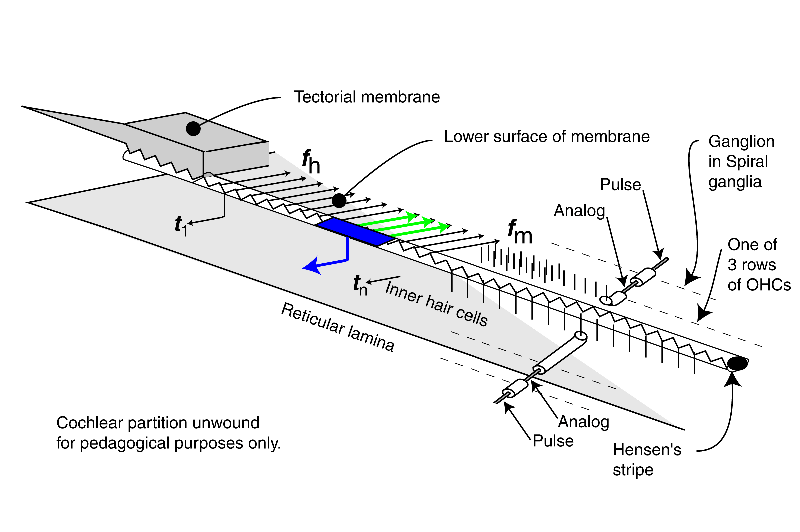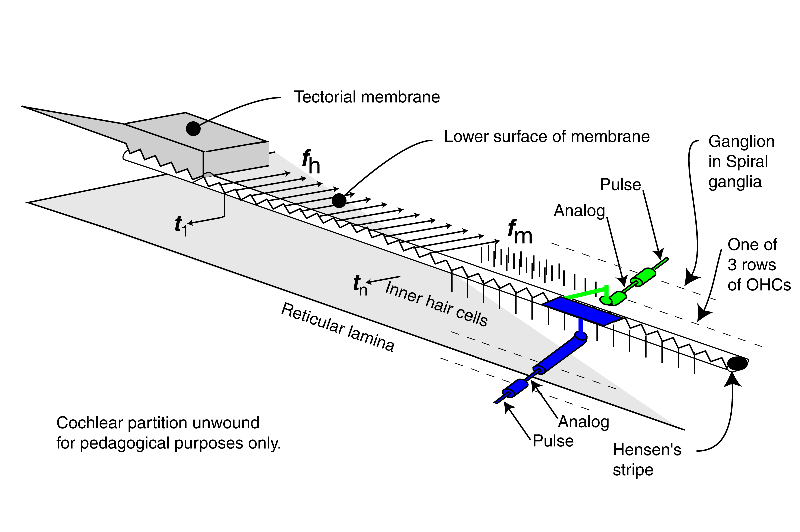
This animation introduces the electrolytic (biologically formed electronic) circuits generating the neural signals. It also differentiates between the analog signals (such as the generator potentials at the pedicles of the sensory neuron axons) and the phasic signals generated by and appearing at the pedicles of later projection neurons.
It is impossible to understand the operation of the cochlea without understanding that each human cochlea contains over 28,000 individual electrolytic circuits (the biological equivalent of man-made electronic circuits) packaged within it.
For the scope and top-level description of the SAW-based Electrolytic Theory of Hearing, see page 18 of Chapter 1 of the e-book "Processes of Biological Hearing"
The following material related to the energy extraction mechanism within the cochlea is drawn from Section 4.6 of Chapter 4 of "Processes in Biological Hearing." A draft of this material can be accessed from the Home Page of this website.
The following material related to the electrolytic amplifiers found in the neural portion of hearing is discussed in Chapters 2 & 5 of "Processes in Biological Hearing." A draft of this material can be accessed from the Home Page of this website.
As described in the previous animation, the energy applied to the cochlea is directed to the SAW launcher at one side of the vestibule of the labyrinth (not the scala vestibuli). This energy is transformed into a slow moving surface acoustic wave known specifically as a modified Rayleigh wave. Such a modified Rayleigh wave is a conventional Rayleigh wave that is propagating along a dielectric waveguide that is not a flat surface. This subject will be discussed more fully in the next animation. In this case, the curved region is called Hensen's stripe.
As shown, the active part of the cochlea forms a bilateral transversal filter. It is bilateral but not opposed. In the physical situation, the hair cells are arranged like the toes of a bilateral human. They all point in the same direction.
As noted in the figure, when unwound, the bilateral tramsveral filter is not frequency selective. Only when the filter is curved does it become dispersive and thereby frequency selective, based on the Marcatili Effect.
The basic signal separation task is performed by removing individual frequency components of the total signal passing along Hensen's stripe sequentially. These signals of a distinct frequency consist of a large variety of signals of short but significant duration. The tonal signals contain specific frequency components that are associated with the "voiced" parts of speech and other sounds. These tonal signals are processed by the Outer Hair Cells of the cochlea. While these tonal signals can be considered orthogonal to each other, they need not all be harmonically related. For the subsets of these frequency components may be harmonnically related, there is no requirement that the fundamental frequency component related to these subsets be present.
The temporal signals related to the envelope of the applied energy do not contain individual frequency components lasting for a significant period of time. While many frequency components are present, their brief duration results in an overall spectrum that is very wide, of very low intensity and difficult to record in the laboratory. These transient signals are processed by the Inner Hair Cells of the cochlea.
The energy associated with the high velocity compression wave, associated with a fluid, is converted into a low velocity surface acoustic wave, associated with the gel surface of the tectorial membrane, within the launcher (energy is shown changing from red to blue within the launcher). The low velocity energy then propagates down Hensen's stripe as shown. The energy is initially concentrated in Hensen's stripe as a transverse wave as suggested by the complex sinusoidal wave at the bottom of the stripe. For a short pulse applied to the oval window, the packet of energy proceeds as shown by the blue packets.
A series of Inner Hair Cells (IHC) are shown arrayed along the length of Hensen's stripe. These cells are in mechanical contact with the stripe. The transverse motions of the gel surface of the stripe presses axially on the cilia of each Inner Hair Cell as the energy packet passes. This motion is transferred to the piezoelectric element within the IHC and thereby generates a temporal electrical signal (shown by the blue arrow and labeled tn describing the envelope of the energy in the packet. The process is very energy efficient.
At the same time, the highest frequency energy associated with the packet is redirected across the gel surface of the tectorial membrane (at an angle of about 24 degrees from perpendicular to Hensen's stripe). The energy in each redirected packet (shown by three frequency distinct arrows in green), with a very narrow frequency range determined by the geometry of the cochlea, encounters the cilia of the first of three rows of Outer Hair Cells (OHC). These cilia are also in contact with the gel surface. The transverse motion of the gel due to the energy of each narrow frequency band packet also causes an axial motion of the cilia. A major part of the energy in this band is transferred to the OHC by this motion. That energy is absorbed by a piezoelectric element within the OHC and thereby generates an electrical signal. What energy that is not absorbed proceeds to the second and then the third cilia and OHC where it is absorbed. The overall process is very efficient.
The above process is repeated until virtually all of the energy has been either dispersed and absorbed by the OHC or absorbed by the IHC. High frequency energy and steep leading edges associated with the envelope of the energy are absorbed near the base of the cochlea. Lower frequency energy and less distinct envelopes associated with the initial energy are absorbed near the apex.
The last portion of the animation shows how the signals are processed initially within the neural portion of the IHC and OHC and in the first ganglion cells synapsing with these sensory neurons. The energy is shown being passed to the sensory neurons (IHC in blue and OHC in green) where it is used to create the generator potential at the pedicles of these cells. This generator potential is an analog waveform with a typical amplitude of 5-10 millivolts (mV) from the nominal -84 mV resting potential of the cell. In the simplest case, each sensory neuron is connected to a single ganglion cell via a synapse. A copy of the generator potential is created in the input circuitry of the ganglion neuron. This signal is used to control a monostable oscillator. As the potential of the generator potential changes, a train of phasic action potentials are created that describe the generator potential using a form of encoding known as time-delay modulation (not frequency modulation). The action potentials have an amplitude typically near +100 mV starting from a resting potential of about -140 mV. The time between pulses can be very long or as short as one millisecond in specialized cases. The minimum time interval is usually 4-5 milliseconds.
Go to the next animation in the operational chain. XXX NOT ACTIVE YET
Go to the roadmap of available animation of hearing. XXX NOT ACTIVE YET
Return to the hearing website home page.