The dual path character of the hearing system becomes clear in the Top Level Schematic of hearing. There are multiple "dual paths." First, the bilateral symmetry found in most animals leads to a dual path architecture at the highest level. Second, the separation of the auditory information into frequency and temporal components leads to a dual channel signal configuration leaving each ear.
The bilateral structure of the system provides several important features: redundancy in case of disease or injury, a gross ability to locate an individual acoustic source, and the ability to recognize a variety of nuances associated with acoustic communications.
The dual character of the signaling system provides the ability to achieve rapid source location as well as a finer but slower mechanism for analyzing the presence of multiple sources. Rapid source location is critically important to the survival of the subject. The finer (but slower) source location process is primarily of value in communications. As an example, it allows the simultaneous location of multiple individual instruments playing in a large orchestra.
At least in the higher mammals, the neural system is highly tailored to support the rapid and slow source location processes. The performance of the circuits involved are qualitatively different.
For the scope and top-level description of the SAW-based Dual-Channel Electrolytic Theory of Hearing, see page 18 of Chapter 1 of the e-book "Processes of Biological Hearing"
The following material related to signal manipularion (Stage 4 Operation) of the hearing system is drawn from Chapter 8 of "Processes in Biological Hearing." A draft of this Chapter can be accessed from the Home Page of this website.
The following material related to the electrolytic amplifiers found in the neural portion of hearing is discussed in Chapters 2 & 6 of "Processes in Biological Hearing." A draft of this material can be accessed from the Home Page of this website.
Some of the nuclei in the following animation are shown displaced to the right for convenience. In fact, the system is highly symmetrical to maintain equal time delays for signals from the two ears.
Each of the nuclei named in the following animation contains at least 4 million individual neural circuits. Their circuit complexity is far beyond our current knowledge. Even the description of the major circuit paths is far from complete. These paths are responsible for performing many parallel tasks.
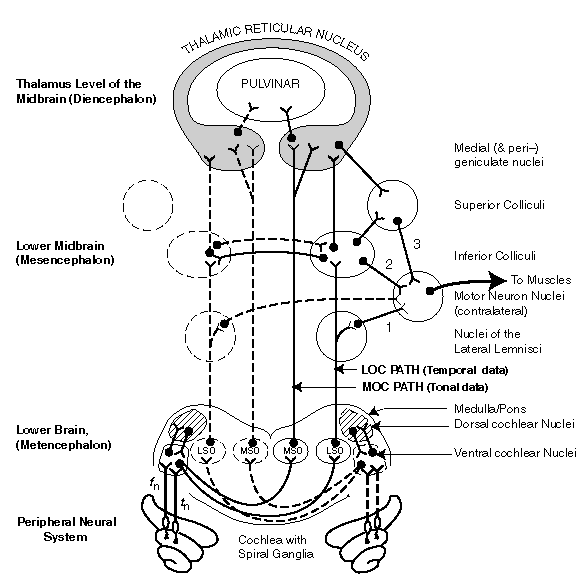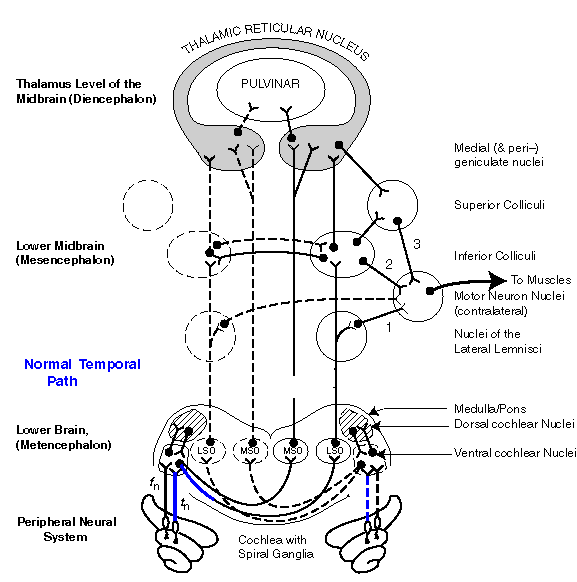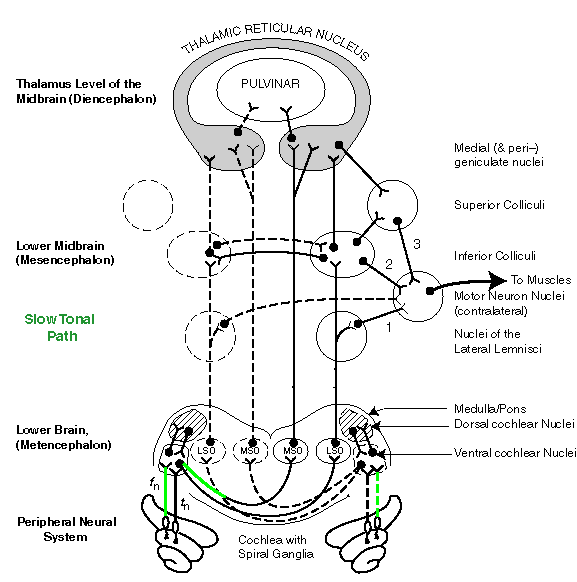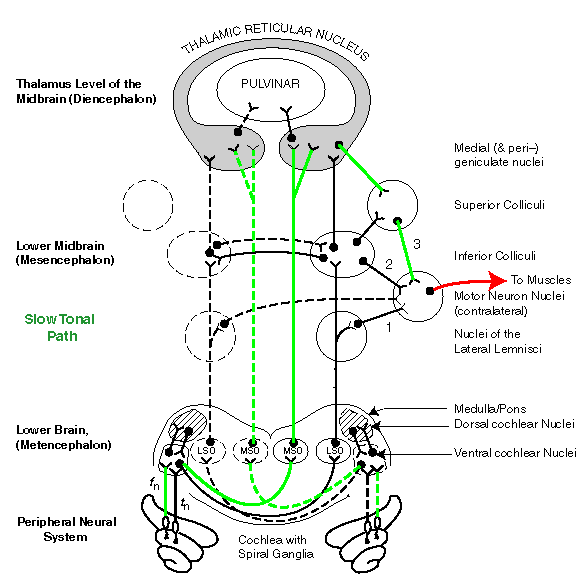
Hearing, like vision, operates in at least five different signal processing modes. The most global purposes supported by these modes are protection of the animal (the Alarm Mode) and awareness of the animals environment (the Awareness Mode). The latter mode includes a major component shared by most animals, intra-species communications or speech. The primary tasks of the Alarm Mode are source location and source identification. Source location will be described first.
Three separate signal paths are used to perform source-location in the higher mammals. The paths converge on the Motor Neuron Nuclei from three different intermediate locations as shown by the numbers 1, 2 & 3 in the figure below. Path 1 is the shortest and least delayed temporal-based Alarm MOde signal path. Path 2 is the normal temporal-based Alarm Mode signal path. Path 3 is a significantly slower frequency-based Awareness Mode signal path.
Path 1, the LOC (temporal) neural paths from the cochlea to the Motor Neuron Nuclei (MNN) via the lateral lemnisci , are believed to be the broadest bandwidth neural circuits in the body. These paths are also as short as possible and skirt much of the signal processing described above. This Alarm Mode path is critically important to the avoidance of danger. At least some of the neurons associated with this path are believed to operate at rates above 500 pulses per second in-vivo.
Path 1, by activating the skeletal muscles, the ocular muscles and the eyelid muscles is a classic example, like the knee jerk, of a reflex arc. This path causes the animal to "turn and duck" before it has determined the character of the threat. In humans, the characteristic response issued somewhat later is, "what was that."
Path 2, the LOC (temporal) neural paths from the cochlea to the MNN via the Inferior Colliculi, is the normal "fast" source-location path . The time difference between the signals received by the two ears, and the shape of the energy envelopes associated with those signals are evaluated. The appropriate MNN are activated, including those of the eyes, and a message is forwarded to the higher brain centers estimating the character of the source.
Path 3, the MOC (frequency-based) neural path from the cochlea to the MNN via the elements of the thalamic reticular nucleus (TRN), is the slowest process for determining source-location. It relies on phase information at frequencies below the integration frequency of the sensory neurons, typically 500 Hertz. Delays greater than five milliseconds are associated with this processing path. The TRN also sends its evaluation of the signals to the parietal lobe of the cerebral cortex for review by the frontal lobe.
Path 3 is not effective at frequencies significantly above the integration frequency. At these frequencies, path 3 only delivers the signature of the stimulus, basically its intensity. Specific frequency or phase information is not provided for these frequencies over this path.
The paths proceeding from the thalamus to the cerebral cortex are not shown in this figure. They deal with the separate and distinct stage described as cognition (Stage 5).
The following animation will describe the three signaling paths described above, the two temporal-path based paths and the single frequency-based path. While shown as single paths, these are "nerve" or "commissure" paths consisting of a great many individual neurons. Those neurons sharing a common processing task are grouped together into these nerves. In the animation, the source generates a signal in the left cochlea first, and this signal proceeds up the contralateral nervous system path. The signal reaches the right ear later and travels up the contralateral path as shown.
Path 1 is illustrated by the blue lines in the first part of the animation. Note the muscle signal is generated before any signal arrives at the TRN and little if any signal reaches the Inferior Colliculi.
Path 2, the normal temporal process of source-location is then shown by the slower moving blue lines in the second phase of the animation. Here the data from both ears is correlated in the Inferior Colliculi and passed to the Motor Neuron Nuclei. Little signal reaches the TRN before the muscles respond. However, data continues to reach the TRN and evaluations are made. These can be used to perform a second order correction on the estimated source location, and to provide an evaluation to the parietal lobe of the cerebral cortex.
Finally, the slow frequency-based source-location signal path, path 3, is shown by the green lines. The signals pass up the nervous system chain to the thalamus where they are correlated based on phase and a signal is generated that can be passed to the Motor Neuron Nuclei by way of the Superior and/or Inferior Colliculus. Since the correlation occurs within the thalamus, it is not necessary to send signals down through both the left and right paths to the Superior Colliculi.
In reality, all of the signals associated with these paths originate from the same external stimlus. The only significant time difference between their starting points is due to the external field geometry and shadowing (or refraction and reflection) by the head and body of the subject.
Go to the next animation in the operational chain. XXX NOT ACTIVE YET
Go to the roadmap of available animation of hearing. XXX NOT ACTIVE YET
Return to the website home page.