The biosonar of the bottlenose dolphin is a very sophisticated high performance system that continues to outperform any man-made system within its operating regime. It does this by employing an array of techniques not fully exploited outside of the biological sphere. While it is believed that all members of Cetacea (whales and dolphins) have at least some echolocation capability, that of the bottlenose appears the most optimized.
The capabilities of this biosonar echolocation system have not been discussed in a comprehensive but detailed report in the technical literture. The system employs a very sophisticated signal generating and receiving system and an equally sophisticated information extraction system. The attached case study, "Dolphin Biosonar Echolocation" will explore many of the features of the system at a level not previously documented.
The phonic and aural systems of the bottlenose dolphin have evolved well beyond that of other chordates, including the bat.
The remainder of this page will provide a brief synopsis of that case study. Only a few references will appear here. The extensive references will be found in the case study.
The order Cetacea (whales, porpoises and dolphins) is well known for its broad use of its unique underwater capability in active echolocation and communications. The capability appears to be particularly well developed in the dolphin, suborder Odontoceti, and particularly in the family of dolphinsTursiops . This study will focus on the most widely studied member of this family, the well known bottlenose dolphin, Tursiops truncatus.
The dolphins are very gregarious, carnivorous, pack animals. The dolphins use their combined vocal and hearing systems so broadly, and their repertoire of signals is so wide, the dolphins are known as "sea canaries." While the dolphins are entirely aquatic animals, their visual and hearing capabilities have evolved to perform in either the aquatic or terrestrial environment to a remarkable degree. They have been known to recognize individual people they know on shore at distances of over 100 .
For echolocation, the bottlenose dolphin is equipped to use two distinct types of biosonar. They are known to use an impulse-type (click-type) biosonar for high precision echolocation, and probably imaging, of targets within 100 meters. Unfortunately, to measure this capabiility, the dolphins have been taught to whistle as a communication with their trainer
The dolphins are quite capable of using their swept continuous tone (whistle-based)capabilities to echolocate on targets with less precision out to about 600 meters. This biosonar offers an intrinsic range and range rate (Doppler) capability but offers a lower angular resolution because of its lower frequency.
In toto, the sonar of the bottlenose dolphin is considerably more sophisticated than any current man-made sonar in the world. It rivals the most advanced airborne radars available today. Merely listing the most obvious properties of this biological sonar (biosonar) attests to its fantastic capabilities. It is fundamentally a multi-band, multimode (including Doppler detection), frequency-hopping, steerable beam, binaural receiver, camouflage penetrating, single-pulse (when required) system with properties at least as sophisticated as the latest stealth fighter plane, the F-117, and latest stealth bomber, the B-2. It is not known how many targets the dolphin can track simultaneously. However, the capability is probably similar to the above man-made systems.
The dolphins, and other members of Cetacea, have evolved their acoustic echolocation capabilities (like the bats) from the generic mammalian system. However, because of their aquatic environment, the dolphins have replaced the morphological structure commonly known as the outer ear (the pinna) by an entirely different structure consisting of a conformal variable-density lens-type receiving system (antenna) that provides excellent forward and omnidirectional performance without slowing the swimming velocity of the animal.
Similarly, the dolphins have replaced sound generation by means of the larynx and the open mouth (oral cavity) with a capability shared between a relocated larynx and a pair of additional phonic lips located within the nasal cavity connecting to the blowhole. They have also introduced an air reuse system that allows them to pass the same air back and forth from special sacs associated with the nasal cavities. As a result, they are able to generate acoustic sounds while underwater without expelling air.
To optimize its performance in its environmental niche, the dolphin has evolved a pneumatic system that is entirely separate from its gastronomic system. It can echolocate and swallow its food simultatneously. The bottlenose dolphin does not emit sounds through its mouth (contrary to the impression left by "Flipper" on the old television series). Those emitted into the terrestrial environment issue from its blowhole, a feature of the extensively modified laryngeal and nasal system of dolphin. Equally interesting, bottlenose the dolphin swallows its food whole (rather than chew with its lower jaw which has adapted into a fragile structure).
Even at this late date, the methods used to create acoustic energy for echolocation purposes are not completely understood. Hoever, it is safe to say that most of the data published before 1995 is archaic and should not be relied upon. The recent introduction of X-ray based computer augmented tomography and magnetic resonance imaging in the medical field has led to major advances in the understanding of the dolphin ehcholocation system. The transmitting system is now known to consist of three different sound generating systems coordinated with a large variable-density lens-type beam former (antenna) known as the melon. It is this melon that sits above the beak of the dolphin in a conformal dome and gives the dolphin's head its distinctive shape.
The conformal arrangement of the transmitting and dual aperture receiving system of the dolphin echolocation system can be seen in the following figure.

A profile-view X-ray can indicate the character of the echolocation system in the bottlenose dolphin. However, a three dimensional recreation by computer-assisted tomography (CT) provides much more information because the soft tissue plays a major role in the operation of the system. A difficulty in interpreting these pictures is the fact that the animal is not designed to be out of water. As a result, some of the soft features may be distorted by gravity. Of greatere importance, some of the soft material forming the acoustic lenses appear to be under muscular control (i. e., they can be reshaped as will by the animal). This capability suggests much more study will be needed to understand the operation of the system under various operating modes created by the muscles. The change in the shape of the conformal receiving antenna is easily seen by comparing pictures of the rear portions of the lower jaw of the bottlenose dolphin.
The sound generation capability of the dolphin can be summarized using the following picture.
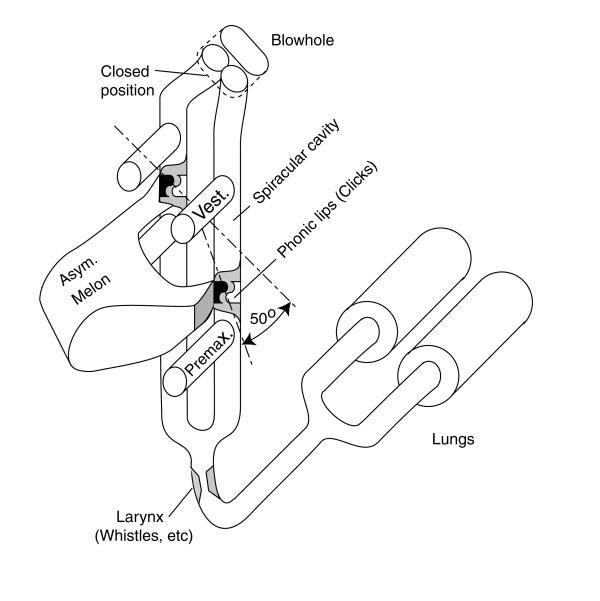
The bottlenose dolphin is capable of producing a wide variety of sounds having intrinsically different spectral structures. A broad series of barks, chatters and squeaks are producible, within the range of human hearing, in both air and water. They can also produce two characteristic sounds in the water. One is a series of one or more short impulses of sound. The fundamental frequency of these pulses is in the 30-150 kHz region. The other is a swept continuous tone. In the latter case, the fundamental tone of the signal is in the 4-20 kHz region. It has also been reported multiple times that the dolphin can generate two separate signals either simultaneously or in rapid sequence. Such performance would suggest the two nasal passages can operate independently in sound production.
This system must be interpreted under a variety of conditions. It is likely that the system operates differently at different depths due to the pressure associated with that depth. Near the surface, it appears the larynx is capable of producing a variety of sounds, not functionally unlike the human, including using the blowhole as an orifice instead of the mouth when the animal is on the surface. This would account for the great range of sounds produced. However, at great depth, it appears the larynx closes and forms a pressure barrier between the lungs and the nasal passages. Differences as great as four atmospheres may occur across this barrier.
The dolphin is able to create a swept continuous tone whistle while under water. It is yet to be shown whether this sound is generated at the larynx or the phonic lips. It is also to be shown whether the animal can produce the swept tone at great depths. If it can, pressure profiles would suggest the swept continuous tone was formed by the phonic lips.
The phonic lips are almost certainly the source of the impulse type signals called clicks produced by the dolphin.
As noted in the figure, the two pairs of phonic lips are displaced in the vertical direction. This suggests the energy from these sources is applied to the melon (acting as an acoustic lens) at different locations. As a result, each source would provide an acoustic beam in the far field with an axis pointing in a different direction. The radiating field pattern of the bottlenose dolphin has been measured and is known to be very complex and to change rapidly with time. In the nominal case, the two pairs of lips could be fired in rapid sequence and generate two radiating beams separated vertically by a nominal ten degrees. If the auditory system can compare the returns from these two pulses, the vertical position of any target within this field angle could be determined with considerably accuracty.
Through evolution, the bottlenose dolphin has abandoned the auditory meatus and external ear (pinna) common to terrestrial mammals and based on a horn "antenna." It has implemented an alternate "antenna" of the lens-type adjacent to each side of the rear of the lower jaw. This form is necessary to satisfy hydrodynamic streamlining requirements while maintaining excellent forward looking acoustic reception.
The form of the system is shown in profile view below.
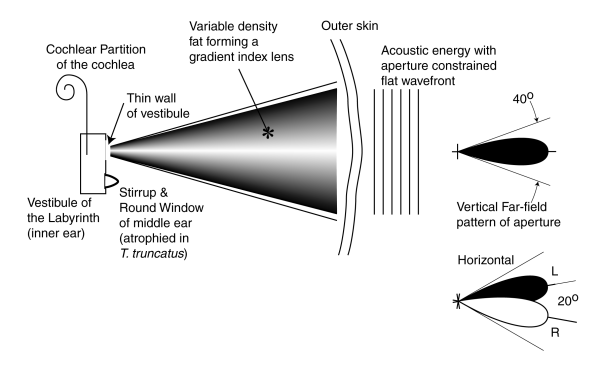
Rather than introduce acoustic signals into the vestibule of the labyrinth via the oval window as in terrestrial mammals, for transfer to Hensen's stripe in the cochlear partition of the cochlea, the energy is introduced through a thin section of the vestibule wall as shown. The energy is brought to the "dolphin window" of the vestibule from the forward facing jowel of the animal, as shown in the above picture, by the fatty material found in this area. This fatty material is known to exhibit a gradient in its density. The fatty material is used to form a gradient density lens-type acoustic antenna as shown. The receiving antenna accepts a nearly planar acoustic wave reflected (or originated) by a target in its external field. It concentrates the energy associated with this wave into a more intense wave of greater curvature that it projects through the dolphin window of the vestibule. Once the energy is delivered to the vestibule, it is passed to the cochlea and processed the same as in any other mammal.
The far-field pattern of the gradient lens-type receiver is shown on the right. The pattern is nominally 40 degrees wide and nearly sidelobe free. The angular width of the pattern is determined by the linear dimensions of the aperture and the frequency of the acoustic energy. The two ears are pointed in different directions, plus and minus 10 degrees relative to the saggital plane, in order to achieve stereoscopic "acoustic viewing" ahead of the animal using active sonar principles. This viewing is functionally different from the passive source location capability associated with binaural hearing. It is functionally similar to, and achieved neurologically in the same way as, stereoscopic vision.
A feature of the gradient density lens-type ear is that it can be steered relatively easily by reshaping the fatty material under muscular control. This allows the dolphin to expand the squint angle while making its external surface more hydrodynamic. The result is a wider area of acoustic search in the horizontal plane while achieving maximum efficiency in swimming velocity. Alternately, the squint angle can be reduced when higher angular resolution is required in a narrower region ahead of the animal.
The azimuth information available from taking the intensity difference between the echos from targets from a single high frequency pulse by the binaural ears and the elevation information available from taking the intensity difference between the echos from targets illuminated by alternate pulses from the bivocal system can be combined to predict the capability of the bottlenose dolphin.
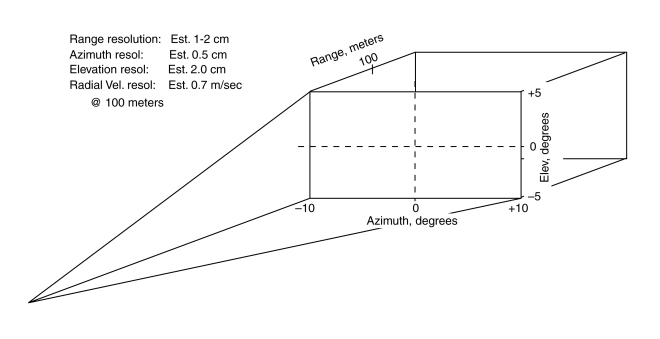
The figure is drawn to represent the nominal squint angle of the ears of 20 degrees in the horizontal plane and the nominal squint angle of the vocalization system of 10 degrees in the vertical plane. The range resolution of the system has not yet been determined with any accuracy.
Again, the bottlenose dolphin follows the neurological plan of other mammals. However, it has undergone significantly different adaptations to this system. The allocation of volume to the portions of the brain greatly favors the paleocortex voer the cerebral cortex in the dolphin. Bullock & Gurevich have studied the dolphin brain closely. They note the size of the medial geniculate is about seven times that in humans, the inferior colliculus is about 12 times its human analog, and the nucleus of the lateral lemniscus is over 250 times as large as the equivalent structure in humans. The ventral cochlear nucleus is also massive compared to the human analog. These are all structures within the paleocortex or "old brain" as opposed to the neocortex or cerebral cortex. As in all mammals, each of these structures is paired.
The animal employs a precision echolocation system (PES) that is very analogous to the precision optical system (POS)found in it and other mammals. Its job is to determine the range and angular direction to a target relative to the animal. The majority of the PES is found in the paleocortex of the brain, along with the POS.
The conformal outer ears deliver the acoustic energy directly to the vestibule associated with the labyrinth and inner ear. The middle ear is not needed as an air-to-fluid impedance transformer since the animal lives in a fluid environment.
The cochlea of the dolphin is almost identical in size and arrangement of the inner and outer hair cells as in the human. It also operates in the same way. The variable curvature of the cochlea disperses the acoustic energy as a function of frequency. This process of dispersion avoids the need for any resonant circuits within the cochlea as often hypothesized. As in the human cochlea, two separate signals streams are generated, one based on the temporal or envelope characteristics of the applied signal using the inner hair cells and one based on the tonal characteristics of the signal using the outer hair cells.
The method by which the acoustic energy applied to the tonal neurons of the cochlea is folded into a multidimensional array in the correlator of the perigeniculate nucleus is illustrated below. The information within the correlator is sensed sensing neurons arranged vertically. Their outputs are transferred to another stage of signal manipulation within the pulvinar of the thalamus as shown at upper right.
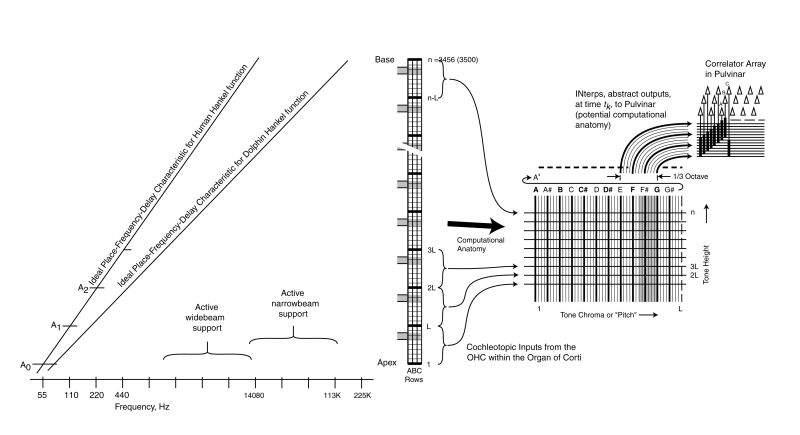
As in some of the neurons emanating from the spiral ganglia associated with the human cochlea, some of the neurons are able to support bandwidths up to between 600 and 1000 Hz. These neural paths are used for precise range determination within the inferior colliculus and the lateral lemniscus.
The massive size of the lateral lemniscus in the dolphin, compared to the terrestrial mammals (it is at best vestigial in humans), suggests it is the seat of signal manipulation related to active echolocation in the dolphin. Similarly, the adjacent inferior colliculus appears to be the seat of passive source location, as it is in other mammals.
As in the human, the other neurons emanating from the spiral ganglia are typically limited to bandwidths of about 250 Hz. These neurons provide accurate place information concerning the energy dispersed in the cochlea but contain no actual frequency components related to the stimulus.
The information extraction circuits of the lateral lemniscus, the acoustic perigeniculate nucleus (the precision portion of the medial geniculate nucleus) use very sophisticated associative correlators to extract multi-dimensional information from the data defining the target scene. These circuits are virtually identical to those used in human vision as well as the vision of other higher mammals.
Because of the similarity of the neural circuitry, it is quite likely that the bottlenose dolphin "sees" an image of the acoustic target space ahead of him/her very similar to the image of visual space he shares with other species. These two images may actually be merged in the higher perceptual circuitry of the dolphin but we have no way of determining whether this possibility is realized at this time. The process would be very similar to the merging of the low spatial resolution peripheral field of vision with the high resolution field of vision provided by the fovea in humans.
The recognized capabilities of the bottlenose dolphin vocal system appear equal to those of the human with respect to interspecies communications. Similarly, the binaural auditory system is equal to that of the human except for its extended high frequency capability. The combined high freqency vocalization and hearing capabililties of the bottlenose dolphin are unequalled in the animal kingdom.
The overall system described above can be used in a variety of one-way and two-way operating modes. The ability of bottlenose dolphins to hear and respond to acoustic signals, within the frequency range of their ability to reproduce, considerably exceeds 200 meters. Best estimates are that the swept continuous tone sonar can locate multiple objects simultaneously to within 10-20 cm out to ranges of 200 meters. This system also provides an excellent Doppler frequency measurement capability. This capability provides the dolphin a relative velocity estimate for a target between two and fifty kilometers/hour with a precision on the order of two kilometers/hour. Well documented experiments have shown the pulse-based acoustic system can locate objects out to 100 meters. While few tests have been performed using multiple targets, the estimate is that they can locate multiple targets with a precision of 10-20 cm out to 100 meters.
Analysis of the overall auditory system of the dolphin suggests it "sees" in the acoustic range with a fidelity equal to that of humans in the visual range (except for the degree of spatial detail). This capability is three-dimensional in character and is achieved with auditory neural ciruitry that is virtually identical to that used in the visual system of themselves and the higher primates. The dolphin has the added ability to measure the depths of targets acoustically as if they were translucent in the visual regime. Interestingly, the swim bladders of other fish and the lungs of other mammals appear as very high contrast mirrors. This feature can considerably increase the detection range of the echolocation system.
Read the Case Study: "Dolphin Biosonar Echolocation"
Return to the hearing website home page.
Bullock, T. & Gurevich, V. (1979) Soviet literature on the nervous system and psychobiology of cetacea Internat Rev Neurobiol vol 21, pp 47-127