
The operation of the cochlea is totally dependent on the Marcatili Effect for frequency discrimination. The effect is widely recognized and utilized within the fiber optics community. The Marcatili Equation contains an argument that is the product of the curvature of the cochlea and the frequency of the dispersed signal.
The cochlea of mammals, birds and reptiles (particularly within the Organ of Corti) all exhibit a characteristic form best represented by an exponential helix. While the bird cochlea is frequently described as only slightly bent morphologically, closer histological examination shows its active structure to be bent about 90 degrees.
The following figure shows the generic form of the cochlea within the active segment known as the Organ of Corti.
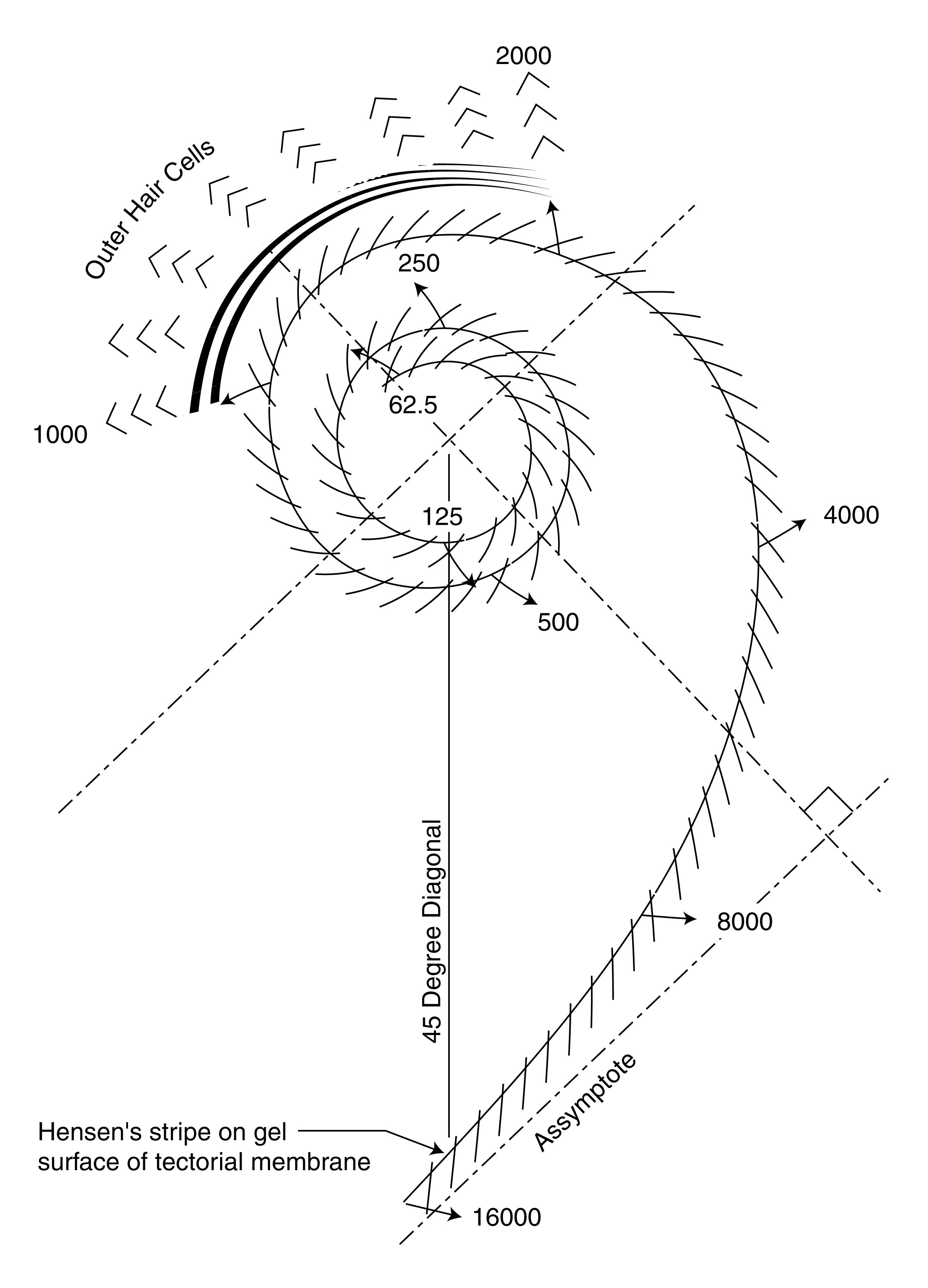
The acoustic energy flows along Hensen's stripe in the form of a surface acoustic wave known as a modified Rayleigh Wave. The energy is dispersed as a function of the curvature (mathematically distinct from the local radius at the point of dispersion) of the stripe. The energy is directed toward the Outer Hair Cells that are present in their typical echelon arrangement. This arrangement assures maximum absorption of the acoustic energy by the appropriate hair cells.
Hensen's stripe differs from a mathematically perfect Henckel Function (a 2-dimensional Bessel Function) due to packaging constraints at low frequencies and ecological requirements among the echolocating species of bats and dolphins at high frequencies. The variations in curvatures within the cochlea of various bat and dolphin species have been documented.
The remarkable attenuation of auditory signals at points along the cochlea beyond the dispersion frequency demonstrates the use of the Marcatili Effect in hearing. No other mechanism is known that can provide such a high degree of attenuation. The following figure compares the attenuation obtainable by a simple resistor-capacitor network, a tuned resistor-inductor-capacitor network with a quality factor (Q) of 25 and the attenuation attributable to the Marcatili Effect alone.
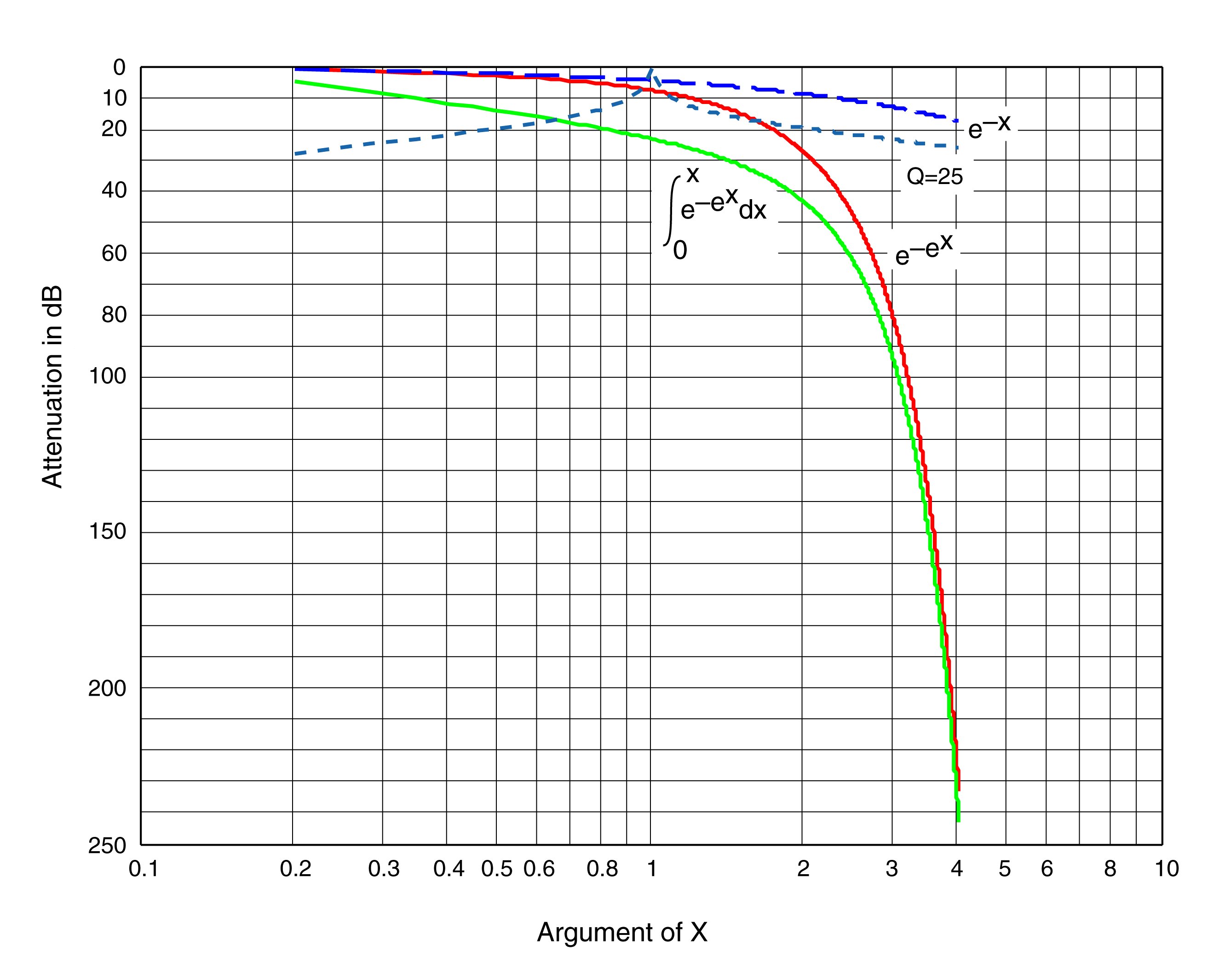
An attenuation exceeding 100 dB is not found in man-made circuitry, even those using solid state physical devices. However, attenuations exceeding 200 dB are frequently measured in cochlear research. Similarly, a rate of attenuation (the slope of the attenuation beyond cutoff) of 180 dB/decade or higher is typically measured in cochlear research.
The Oto-acoustic emission (OAE) performance within human and other mammalian species has caused great confusion among researchers because of the absence of certain sidebands expected to be generated by non linear mixing of specific tonal frequencies. The following figure shows why some of these sidebands, particularly at higher frequencies are suppressed within the retrograde signals within the Organ of Corti.
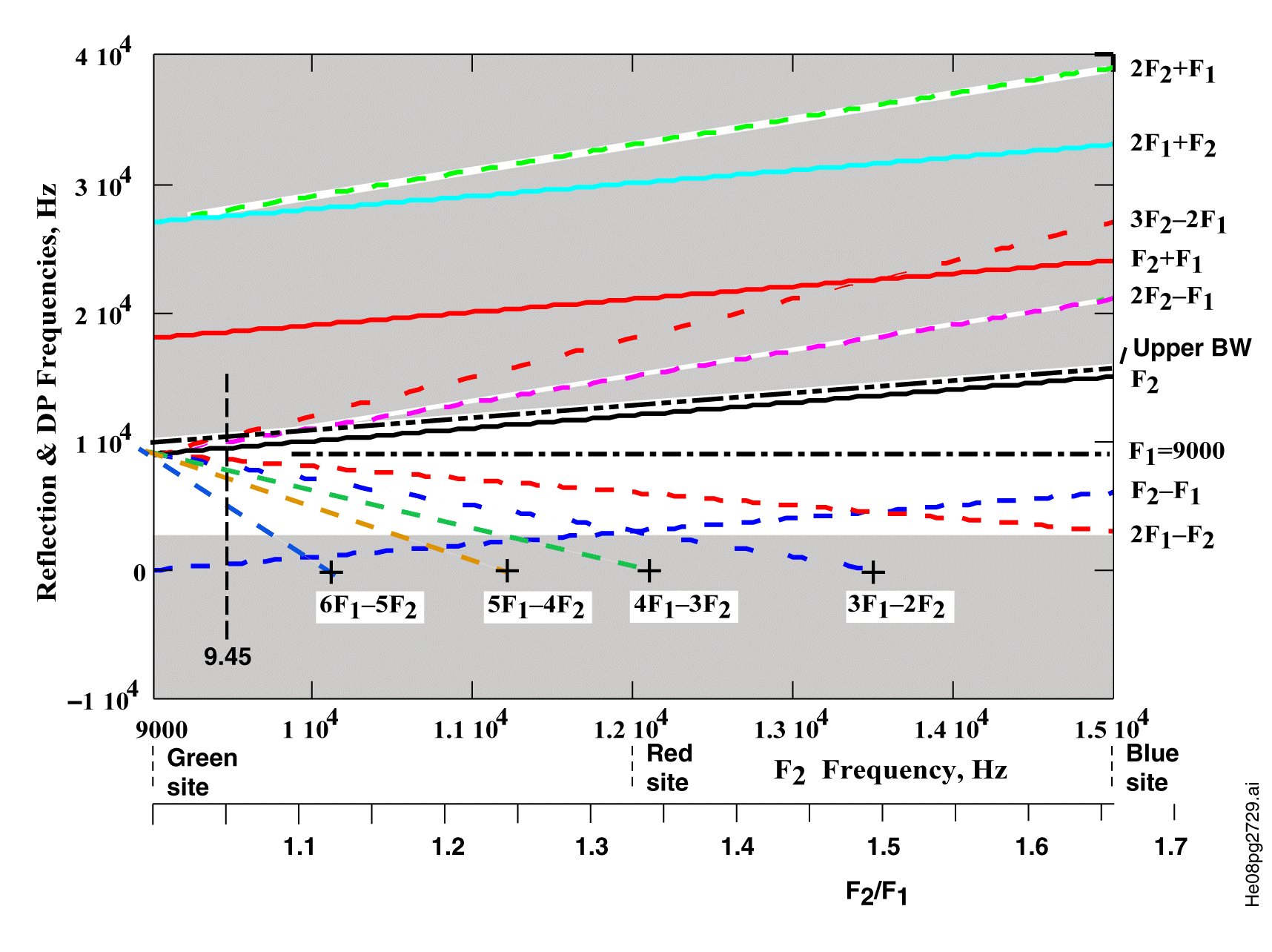
Distortion products generated in the shaded areas are greatly attenuated as the signals attempt to progress retrograde across the areas of very high attenuation due to the Marcatili Effect within the Organ of Corti. Note the wedge shaped area of minimal attenuation. No upper sideband distortion products are observed by a microphone in the outer ear canal. Only a few distortion products at lower frequencies are observed. More distortion products due to oto-acoustic excitation (DPOAE) are observed at smaller ratios of F2/F1 but the experimental instrumentation is more demanding in this region. For F2/F1 greater than 1.35, only the 2F1 - F2 sideband is typically observed.
No other theory of hearing has successfully explained the above characteristics in either qualitative or quantitative terms. A more complete discussion supporting the above assertions can be downloaded. It is in turn supported by more detailed presentations in Chapter 4 and Chapter 9 of the Hearing: A 21st Century Paradigm listed in the reference section.
Return to the Hearing website home page.
Return to the Neural Concepts website home page.
Material presented by James T. Fulton
Fulton, J. (2008) Hearing: A 21st Century Paradigm. Bloomington, IN: Trafford Publishing Chapters 4 & 9 Also at
http://neuronresearch.net/hearing/pdf/4Physiology.pdf and
http://neuronresearch.net/hearing/pdf/9Performance.pdf
Marcatili, E. (1969) Bends in optical dielectric waveguides BSTJ vol 48, pp 2103-2132
Marcatili, E. & Miller, S. (1969) Improved relations describing directional control in electromagnetic wave guidance BSTJ vol 48, pp 2161- 2181
Evans, E. (1975) Cochlear nerve and cochlear nucleus In Keidel, W. & Neff, W. eds. Handbook of Sensory Physiology: Auditory Systems, volume 5.2, pages 1108 Berlin: Springer pg 15
Fay, R. (1990) Structure and function in sound discrimination among vertebrates In Webster, D. Fay, R. & Popper, A. eds. The Evolutionary Biology of Hearing. NY: Springer-Verlag. Chap 14, pg
He, W. Fridberger, A. Porsov, E. Grosh, K. & Ren, T. (2008) Reverse wave propagation in the cochlea PNAS vol 105(7), pp 2729-2733